Articles
The camerate crinoid Scyphocrinites Zenker in the Upper Silurian or Lower Devonian of New Brunswick, Canada
ABSTRACT
The middle Paleozoic Scyphocrinites Zenker has a distal attachment modified into a globular flotation structure and, uniquely for a crinoid, joined the obligate plankton. Such a flotation structure has been found in the Indian Point Formation (Pridolian to Lochkovian) of Flatlands, northern New Brunswick. It is most likely of Pridolian (Late Silurian) age based on the primitive morphology. The identification is confirmed by the globular gross morphology, multi-plated calcite structure, age and similarity to coeval fossils from Cornwall, southwestern England.
RÉSUMÉ
Le Paléozoïque moyen Scyphocrinites Zenker a une attache distale modifié dans une structure de flottaison globulaire et, uniquement pour un crinoid, rejoint le plancton obligatoire. Une telle structure de flottaison a été trouvée dans le point de Formation d’Indian (Pridolien à Lochkovien) de Flatlands, le nord du Nouveau-Brunswick. Il est fort probable Pridolien (Silurien supérieur) sur la base de la morphologie primitive. Cette identification est confirmée par la morphologie globulaire brut, la structure de la calcite multi-plaqué, de l’âge et de la similarité des fossiles contemporaines de Cornwall, sud-ouest de l’Angleterre.
[Traduit par la redaction]
INTRODUCTION
1 Stalked crinoids are typically part of the modern deepwater benthos, but our appreciation of their life orientation and other ecological/paleoecological preferences has been unduly influenced by Nineteenth Century interpretations rather than modern observations (Donovan 2011a). For example, it comes as a surprise to many that some modern stalked crinoids may crawl with their arms (Baumiller and Messing 2007), and the stalkless comatulids can use the same structures for walking or swimming (Meyer and Macurda 1977). They are thus benthic, but certainly not sessile. In the ancient past, some crinoids attached to floating objects such as logs and pursued a pseudoplanktonic lifestyle (Simms 1986; Donovan 2011b); alternatively, in the case of the middle Paleozoic genus Scyphocrinites Zenker, the roots were modified to form a globular flotation device, the lobolith, which made these crinoids true members of the ocean plankton. No other crinoid has ever evolved such a bizarre distal structure (Fig. 1).
 Display large image of Figure 1
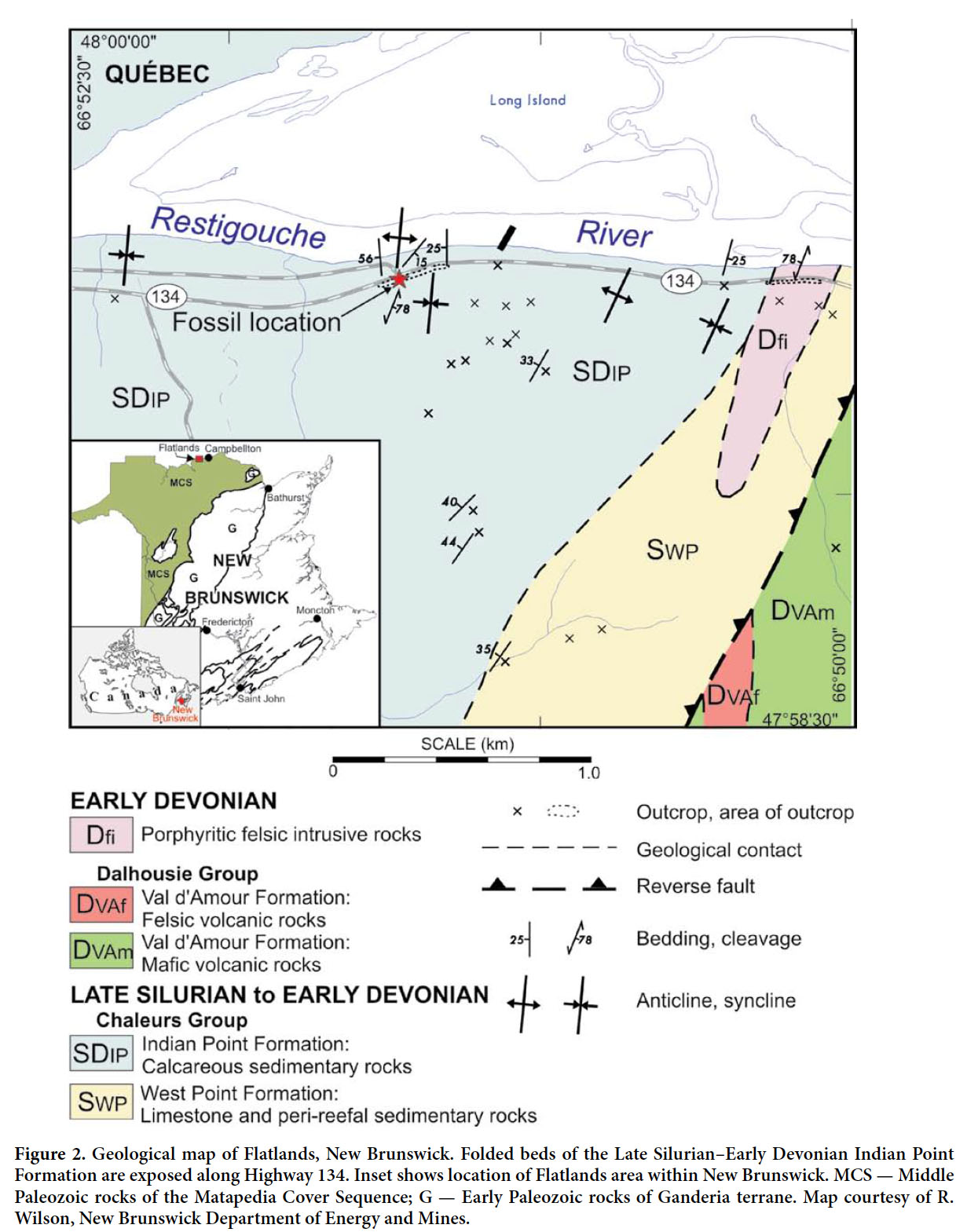
Display large image of Figure 12 The planktonic habit of Scyphocrinites led to it being a facies-crossing taxon in the sedimentary record, distinguishing it from most other Paleozoic crinoids. Further, it has a limited stratigraphic range, occurring in strata that span the Upper Silurian (Pridolian) to Lower Devonian (Lochkovian) boundary, so it is a rare crinoid that can contribute to international biostratigraphic correlation. Its occurrence varies from the gregarious (e.g., Lee 2005) to the diffuse, the latter being the case for the example that we record herein from a Silurian-Devonian boundary section at Flatlands, northern New Brunswick (Fig. 2).
3 Terminology used for the crinoid endoskeleton follows Moore et al. (1978) and Ubaghs (1978a). Specimens discussed herein are deposited in the Natural Science Department, New Brunswick Museum, Saint John (specimen numbers prefixed by NBMG) or the Geological Museum, University of New Brunswick, Fredericton (UNB).
 Display large image of Figure 2
Display large image of Figure 2LOCALITY AND HORIZION
4 The fossil was found in a loose block along a road cut of the Indian Point Formation, on the south side of Highway 134 opposite Island View Road, approximately 1 km east of Flatlands, New Brunswick (Fig. 3). Tumbledout blocks of calcareous mudstone from this locality previously yielded specimens of Medusaegraptus, the intact thalli of a noncalcified macroalgal taxon (LoDuca et al. 2013). Additional fossils in the same blocks include scarce brachiopods, trilobites and crinoid fragments. No trace fossils are present in the blocks.
5 The Indian Point Formation consists mainly of moderately to strongly calcareous mudstone, fine-grained sandstone and wackestone, and commonly contains fossils of stromatoporoids, brachiopods, trilobites, crinoids and rugose corals (Wilson et al. 2004). Spores and acritarchs (Wilson et al. 2004), as well as brachiopods from the Indian Point Formation in Gaspé, Quebec (Bourque and Lachambre 1980), indicate a Pridolian to middle Lockhovian age. Wilson et al. (2004) reported a U-Pb (zircon) age of 415.0 ± 0.5 Ma for the Squaw Cap Felsite, which intrudes the Indian Point Formation.
 Display large image of Figure 3
Display large image of Figure 36 A shallow nearshore marine setting was postulated by Wilson et al. (2004) based on the presence of terrestrial microfossils mixed into a marine environment dominated by brachiopods, crinoids, corals, stromatoporoids and trilobites. Although the rarity of cross-laminated or ripple-laminated beds indicates deposition below fair-weather wave base, Wilson et al. (2004) suggested that water depth may have been only a few meters, as seafloor topography related to reefs in the West Point Formation may have acted to baffle wave and current energy. This interpretation is consistent with the presence of Medusaegraptus (LoDuca et al. 2013).
SYSTEMATIC PALEONTOLOGY
- Class Crinoidea J.S. Miller 1821
- Subclass Camerata Wachsmuth and Springer 1885
- Order Monobathrida Moore and Laudon 1943
- Family Scyphocrinitidae Jaekel 1918
Genus Scyphocrinites Zenker 1833
7 TYPE SPECIES. Scyphocrinites elegans Zenker 1833, p. 25, by monotypy (Ubaghs 1978b, p. T489).
8 OTHER SPECIES. Webster (2003) listed 15 nominal species of Scyphocrinites (including Camarocrinus Hall 1879) that he regarded as valid.
9 DIAGNOSIS OF LOBOLITH. (After Ubaghs 1978b, p. T489.) “Column ending distally either in small encrusting root or (more typically) in large bulbous chambered structure designated separately as Lobolithus or Camarocrinus with walls of minute irregular plates, having their origin in bifurcations and modifications of innumerable rootbranches; axial canal of stem quinquelobate to sharply quinquestellate.”
10 REMARKS. As noted by Lee (2005), in light of the pivotal research of Haude (1992), loboliths of Scyphocrinites fall into two morphological groups of broadly similar gross morphology. The primitive condition is represented by the so-called cirrus (more correctly these are inflexible radices, not flexible cirri; Donovan 1993) loboliths, formed from amalgamated branching “roots”; this group includes the specimen from Flatlands (Fig. 4). Cirrus loboliths had been considered to be mainly limited to the Pridolian (e.g., Haude 1992; Lee 2005, p. 334), but the primitive condition is now known also from the lowermost Lochkovian (Donovan and Lewis 2009). The more robust plated loboliths (Fig. 2) range from the uppermost Pridolian to lowermost Lochkovian.
Crinoids remain poorly known from the Silurian of New Brunswick. Specimens in the NBM are currently being described by the authors (research in progress) and newly collected material, provisionally assigned to columnal morphospecies Cyclocyclicus (col.) geoffnewalli Fearnhead and Donovan 2007, is being described by one of us (SD) and D.G. Keighley (University of New Brunswick).
11 RANGE. Upper Silurian (Pridolian) to Lower Devonian (lowermost Lochkovian), global in distribution (Haude 1992).
Scyphocrinites sp. (Fig. 4)
12 MATERIAL, LOCALITY, AND HORIZON. A single specimen, NBMG 16862, from Flatlands, New Brunswick. Collected from a tumbled-out block along the south side of Highway 134 (Figs. 1–2), opposite Island View Road (NTS Map 21 O/15; 47°59.47’ N; 66°51.47’ W). Indian Point Formation, Pridolian (Upper Silurian) or, less probably, Lockhovian (Lower Devonian); see above. Collected by RFM, September 10, 2012.
 Display large image of Figure 4
Display large image of Figure 413 DESCRIPTION. Irregularly-shaped clast derived from a bed 60–65 mm in thickness, composed of well-lithified, grey, micaceous, fine-grained sandstone, weathering brown. The lobolith is collapsed and preserved as an irregularly rounded area about 132 × 113 mm. The rim of this area is continuous, more or less thickened; more centrally, less than half of the area is covered by articulated echinoderm plates, these being either lost by exfoliation or concealed by sandstone. Echinoderm plates contrast with the brownweathered rock because of their medium to dark grey coloration (Fig. 4). Plates are numerous, rounded in outline and swollen, in lateral contact (where apparent) and forming regular to irregular plate aggregations. Attachment for the column is not preserved and there are no conspecific columnals or pluricolumnals preserved on adjacent bedding surfaces. Other bioclasts on more or less the same bedding plane include solitary rugose corals, brachiopods (including a rhynchonellid) and some poorly preserved crinoid debris. Other, unregistered specimens from the same site, now in the same drawer, include what may be columnals and pluricolumnals of Scyphocrinites sp.
14 REMARKS. Four lines of evidence support our identification of this peculiar specimen as a lobolith of Scyphocrinites. The globular gross morphology is reminiscent of how similarly shaped, weakly sutured structures such as Paleozoic echinoid tests collapse in the rock record (Lewis and Emson 1982). The multi-plated calcite structure is certainly echinodermal. The age of the Indian Point Formation is limited, but includes the entire range of Scyphocrinites (Haude 1992). And these data are supported by the close morphological similarity of NBMG 16862 to other occurrences of multi-plated loboliths (discussed below).
We therefore interpret this specimen as an incomplete, partly collapsed and partly deformed lobolith of Scyphocrinites sp. The mode of preservation is reminiscent of loboliths originally described by Bather (1907) from the Lochkovian of Cornwall, southwestern England. The Cornish specimens are so poorly preserved that they were not figured in Bather’s report and have only been illustrated more recently (Donovan and Lewis 2009, fig. 1; Donovan et al. 2012, text-fig. 77; Donovan and Fearnhead 2014, pl. 15). The principal difference between these occurrences is lithology, as the Cornish specimens are preserved in limestone. This difference in occurrence is not surprising in a crinoid that is planktonic; Simms (1986, pp. 476–477, table 1) demonstrated similar facies-crossing distributions of pseudoplanktonic crinoids from the Jurassic.
Our new record of Scyphocrinites sp. is notable because Silurian and Devonian crinoids remain poorly known from eastern Canada. Previous reports have demonstrated the utility of the crinoids from this region for studies of paleoecology and morphology (Donovan and Pickerill 1995a, b). Part of the reason why crinoids from this area and interval have been ignored is undoubtedly taphonomic. Few of the crinoids known from this region are well preserved, again with the notable exception of material from Anticosti Island, and there is a resistance to expending energy on fragmentary fossils when more complete, aesthetically more pleasing and easier to identify taxa, such as brachiopods, trilobites and corals, are available. This is a common problem with Paleozoic crinoids (see, for example, Donovan 2013) and, in the absence of more complete material, it is only by studying incomplete specimens like NBMG 16862 that our knowledge of their true biodiversity and paleogeography will be completed.
SKD gratefully acknowledges the award of a G.F. Matthew Research Grant for 2013 by the New Brunswick Museum, which facilitated this research in May 2014. Naturalis Biodiversity Center, Leiden, provided further support and further facilitated a research visit to the New Brunswick Museum in May 2014. We thank Fiona Fearnhead (the Natural History Museum, London) and Christopher Paul (University of Bristol) for their positive comments in reviewing the manuscript. Thanks to Reg Wilson, New Brunswick Geological Surveys Branch, Department of Energy and Mines for permission to reproduce Fig. 2.