Notes
Digital compression maps:
an improved method for studying Carboniferous foliage
ABSTRACT
A medullosalean-pteridosperm specimen, 22 cm long, from the Sydney Coalfield, Nova Scotia, Canada, illustrates the advantage of studying the fossilization history of compressed foliage freed from the rock matrix by hydrofluoric acid, as compared to the examination of compressions still in the rock matrix. The image of any freed frond segment of compression foliage that has been reprocessed digitally to represent its original structure is called a compression map. Interpretation of a compression map is reliant on a physicogeochemical model of preservation processes.
RÉSUMÉ
Un spécimen fossile d’une ptéridospermée du genre Medulossa mesurant 22 cm de longueur et excavé dans le bassin houiller de Sydney, en Nouvelle-Écosse (Canada), illustre bien l’avantage d’étudier l’histoire géologique de fossiles de feuilles extraits de la matrice rocheuse au moyen de l’acide fluorhydrique plutôt que d’examiner des compressions qui sont encore dans la matrice. On désigne carte de compression l’image d’un segment de fronde compressé extrait de la matrice dont on a fait l’extrapolation numérique pour voir sa structure originale. L’interprétation d’une carte de compression repose sur un modèle physicogéochimique des processus de préservation.
[Traduit par la redaction]
INTRODUCTION
1 A foliar compression is an analogue of a leaf composed of a cuticle (a biomacropolymer) that envelops vitrinite or some other maceral derived from the leaf mesophyll (Zodrow et al. 2009). As plant fossils, foliar compressions are most frequently encountered in Carboniferous strata (Schopf 1975; Zodrow et al. 2009). Pioneering work by Gothan (1916) and Barthel (1961; 1962) in studying Permocarboniferous pteridospermous cuticles, and subsequent work by others (e.g. Cleal and Zodrow 1989; Šimůnek and Cleal 2011) is mostly based on cuticles extracted from a limited number of compressions per species. This approach has nevertheless been successfully used for systematic revision of some compressed Carboniferous neuropteroids (e.g., Cleal et al. 1990).
2 In this paper, we describe how to prepare and present compressions maps. What can be learned from them about the fossilization process of foliar compressions is presented in terms of four variables: heat; temperature; acidity (pH); and oxidation and reduction (Eh). This is then summarized in a proposed preservation model.
BRIEF DESCRIPTION OF THE SPECIMEN
3 The medullosalean-pteridosperm segment (Fig. 1a) originated from the roof shale of the Lloyd Cove Seam (basal Cantabrian), Sydney Coalfield, Canada (Zodrow 2002, Fig. 1). It is compressed with preserved cuticle as is commonly the case in this roof rock. Some compressions appear thin and show an amber hue that is indicative of the fossilized-cuticle that resulted from natural oxidation (Zodrow 1985; Zodrow and Mastalerz 2009). The frond segment is characterized by rapid pinnule differentiation over six distal ultimate pinnae, from odontopteroid at the top, neuropteroid in the middle, and tripartite at the base. Not knowing the fully developed morphology precludes indisputable taxonomic assignment, though our latest spectrochemical research suggests affinity with Macroneuropteris Cleal et al. 1990 (unpublished data of present authors). Such heterogeneous fronds are not uncommon in the Carboniferous “Neuropterid Galaxy” (see Laveine 1997; Barthel and Amelang 2011).
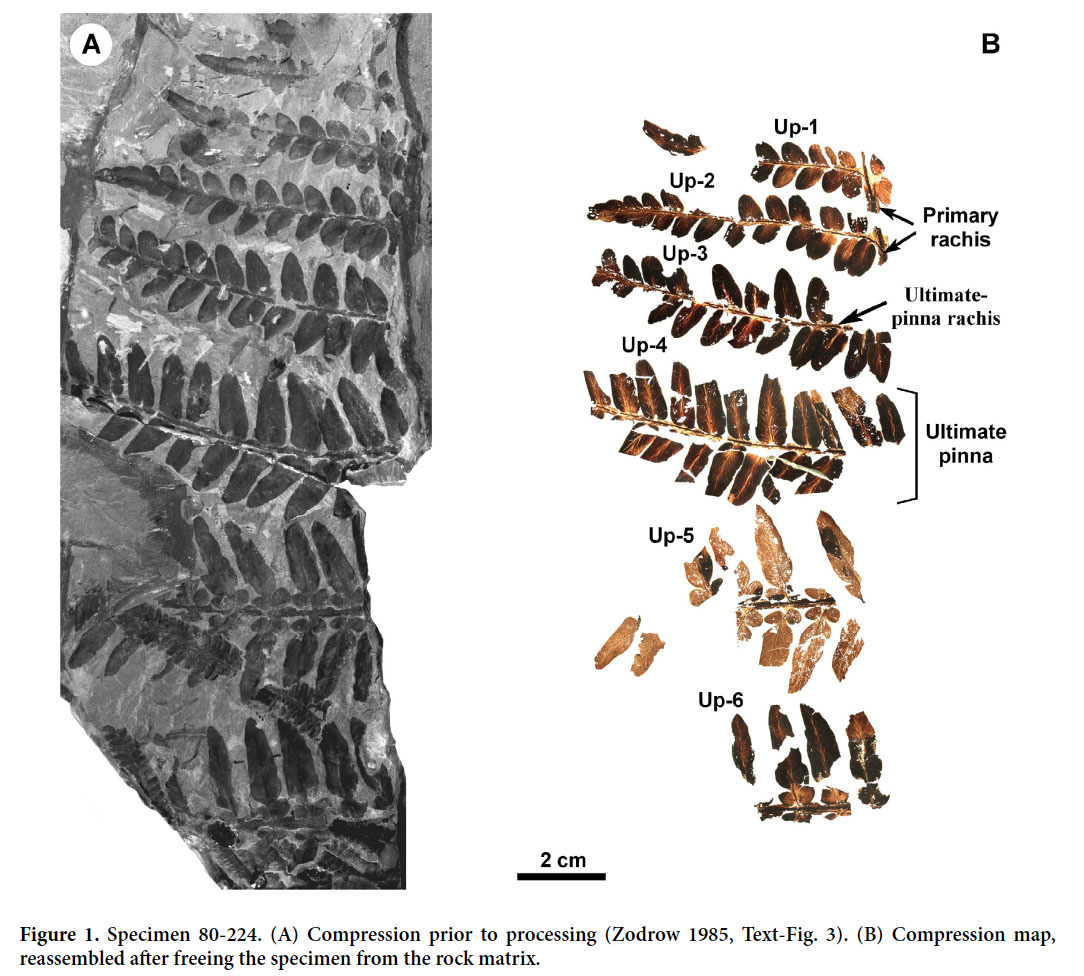 Display large image of Figure 1
Display large image of Figure 1SPECIMEN PREPARATION
4 A compression is not just a “coaly film” but is about 30 µm thick and composed of complex organic structures (mainly aliphatic and aromatic hydrocarbons). Using utmost care, the frond segment is cut with a rock saw into constituent ultimate pinnae, which are digitally photographed. Then, the rock matrix of each pinna is trimmed with the saw to a thickness of < 0.5 mm; this decreases digestion time in hydrofluoric acid (HF) for freeing the compressions. After HF treatment, the pinnae are carefully washed in distilled water for several days to remove water-soluble HF residue. No additional chemical treatment is applied.
5 In the case of our specimen, each of the freed 32 ultimate-pinna fragments was photographed using transmitted light at the same orientation and constant intensity to ensure direct transparency/opacity comparisons. Software-image processors were used to reassemble individual digital photographs (i.e., ultimate-pinnate fragments) to correspond to the original frond structure. This is the compression map (Fig. 1b), which, when compared with the original frond segment, (Fig. 1a), shows a loss of only a few pinnules, notably for Up-5; the overall loss was <10 %.
PREREQUISITES FOR COMPRESSION MAPS
6 The foremost prerequisite in producing a compression map is the availability of contiguous frond segments rather than an interpreted reconstruction from separate specimens; the latter would produce dubious results. Secondly, an understanding of the diagenetic influences on the specimen is critical. Diagenetic factors include: thermal history (Barthel 1961; Hacquebard 1998); pH-Eh redox potential (oxidation-reduction reaction: Krumbein and Garrels 1952; Gautier 1985; Richardson and McSween 1989); bacterial impetus (Zobell 1946); and lithology (summarized in Zodrow et al. 2009). Empirical evidence pointing to favorable preservation conditions at the Lloyd Cove Seam includes the following: (a) entombment of plant fossils in an unaltered clayey-rock matrix of roof rock and seat earth (Deer et al. 1992); (b) favorable vitrinite reflectance (Ro%) ca. 0.65%, volatile matter ca. 36%; and (c) acidic conditions (pH <7 and -Eh; Zodrow and Mastalerz 2009). The last is inferred from the framboidal pyrite associated with the fossilized-cuticles (Zodrow and Mastalerz 2009), which reflects early diagenetic conditions (Birk 1990).
USES OF COMPRESSION MAPS
7 In general, a compression map facilitates detailed physical study, especially of taxonomically important features that are obscured, or not observable, when the fossil was still imbedded in the rock matrix. For example, tangential-lateral veins separated by one or two epidermal cells can be determined as non-meshed veins, critical in distinguishing Linopteris from Reticulopteris (Zodrow and Vasey 1986); and the presence of sparse hairs would be a determining factor in the identification of Macroneuropteris scheuchzeri (Zodrow and others unpublished data). Trichomes that otherwise might have been obliterated by maceration (Gothan 1916) are clearly identified on compression maps, allowing the origin of structural holes on the cuticles to be determined.
8 Visual evaluation of preservation variability is the most significant information obtainable from compression maps as it underpins subsequent interpretations of chemical, palaeoenvironmental, statistical, and cuticular studies. Such evaluation is a function of the degree to which coal (opaque vitrinite) persists in the compression, and thus on the variability of light transmission through the specimen. In an oxidizing medium, be it during diagenesis or by chemical analyses as first done by Stopes (1919), the vitrinite becomes soluble, whereas the transparent biopolyester cuticle remains intact (Zodrow and Mastalerz 2009; Benítez et al. 2004). Thus, the vitrinite and cuticle components of a compressed pinnule are the result of differing reaction paths during diagenesis, processes that are chemically traceable and quantifiable by Fourier transform infrared spectroscopy or by 13 C nuclear magnetic resonance analyses.
9 Interpreted on the basis of differing reaction paths, the present compression map shows influences of natural oxidation that varied in intensity considerably over the small frond segment. For example, the transparent (highly oxidized) “compressions” near the centre, Up-5, and distally at Up-1, contrast markedly with more opaque (less oxidized) compressions elsewhere in the frond segment. Also, midveins are consistently more oxidized than pinnule laminae, but vascular strands in the primary-pinna rachis at Up-1, and generally in the ultimate-pinna rachides, are preserved presumably as original lignin.
10 Results from infrared analyses are in line with these observations, particularly the fact that a decrease in natural oxidation correlates with higher aromaticity and higher condensation of aromatic carbon-nuclei, which principal component analysis confirms. In other words, the degree of transparency correlates with the variable chemistry, the coaly component being compromised as a result of fossilization (D’Angelo et al 2012).
11 From these observations it is clear that plant preservation modes such as compressions, coal balls, and natural-oxidation products that we call fossilized-cuticles, cannot be viewed as isolated phenomena. Rather, they are complex products that formed in given geochemical systems in which preservational gradualism prevailed — preservation modes can imperceptibly overlap or even overprint in a continuous process. The proposed model (Fig. 2) conveys this dynamism by showing that P, T, pH and Eh approach, but are not at, the apices of the tetrahedron. Thus, after burial a leaf can be transformed through stages of compression into a fossilized-cuticle and ultimately graphite, the result that we see depending on the intensity of transformation to the present.
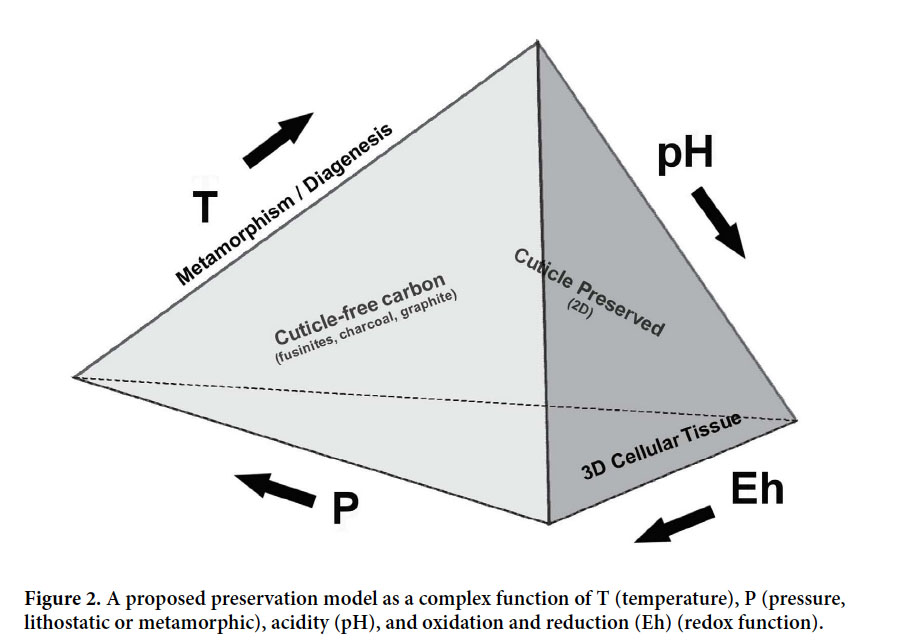 Display large image of Figure 2
Display large image of Figure 212 Although redox conditions for the present specimen have not been investigated, oxidative effects are observable in the laboratory whenever a cuticle is obtained from a compression. Thus, for the present frond segment, we assume that local redox conditions varied. In a future publication we will further address the proposed model.
CONCLUDING REMARKS
13 To our knowledge, no studies have been published that deal with a general theory of plant-fossil preservation, though Krumbein and Garrels (1952) and Berner (1980) provided theoretical approaches in this respect. The digital compression map offers an effective means of visually assessing both the physical attributes and the preservation quality of compressions as a guide for further analytical studies. A significant discovery of our test case is that that, even over a small frond segment, fossilization history is remarkably unstable, probably best explained by local variation in redox potentials.
We thank M. K. Jones, Biology Department, Cape Breton University, for use of microscopes, and M. Barthel, Museum für Naturkunde, Humboldt-Universität zu Berlin, for comments on an earlier manuscript. Journal editor R. A. Fensome and reviewers A. Bashforth, Department of Palaeobiology, Smithsonian Institution and J. H. Calder, Nova Scotia Department of Natural Resources, deserve much credit for critical comments and editorial suggestions that resulted in improving content and presentation of the manuscript; we thank them all cordially.