Articles
The Joggins Fossil Cliffs UNESCO World Heritage site:
a review of recent research
ABSTRACT
The Joggins Fossil Cliffs UNESCO World Heritage Site is a Carboniferous coastal section along the shores of the Cumberland Basin, an extension of Chignecto Bay, itself an arm of the Bay of Fundy, with excellent preservation of biota preserved in their environmental context. The Cliffs provide insight into the Late Carboniferous (Pennsylvanian) world, the most important interval in Earth’s past for the formation of coal. The site has had a long history of scientific research and, while there have been well over 100 publications in over 150 years of research at the Cliffs, discoveries continue and critical questions remain. Recent research (post-1950) falls under one of three categories: general geology; paleobiology; paleoecology. It provides a context for future work at the site. While recent research has made large strides in our understanding of the Late Carboniferous, many questions remain to be studied and resolved, and interest in addressing these issues is clearly not waning. Within the World Heritage Site, we suggest that the uppermost formations (Springhill Mines and Ragged Reef), paleosols, floral and trace fossil taxonomy, and microevolutionary patterns are among the most promising areas for future study.
RÉSUMÉ
Le site du patrimoine mondial de l’UNESCO des falaises fossilifères de Joggins est situé sur une partie du littoral qui date du Carbonifère, sur les rives du bassin de Cumberland, qui est une prolongation de la baie de Chignecto, elle-même un bras de la baie de Fundy. L’endroit offre un excellent milieu de préservation de la faune et de la flore dans leur environnement. Les falaises donnent un aperçu du monde du Carbonifère tardif (Pennsylvanien), soit la période de l’histoire de la terre la plus importante pour la formation du charbon. Ces falaises ont fait depuis longtemps l’objet de travaux de recherche scientifique et plus de 100 publications y ont été consacrées au cours de 150 années de recherche. L’endroit suscite encore des découvertes et il soulève toujours des questions essentielles. Les travaux de recherche récents (depuis les années 1950) se répartissent en trois catégories: géologie générale; paléobiologie; et reconstitution des paléomilieux, en plus d’offrir des avenues pour les futurs travaux qui devraient s’y dérouler. Même si la recherche récente a largement contribué à une meilleure compréhension du Carbonifère tardif, de nombreuses questions demeurent sans réponse et commandent qu’on les étudie et les résolve et il est manifeste que l’intérêt pour ces questions est bien loin de s’amenuiser. Sur ce site du patrimoine mondial, nous soutenons que les futurs sujets d’étude les plus prometteurs seraient les formations supérieures (Springhill Mines et Ragged Reef), les paléosols, la taxonomie florale et des ichnofossiles, ainsi que les paramètres de microévolution.
[Traduit par la redaction]
INTRODUCTION
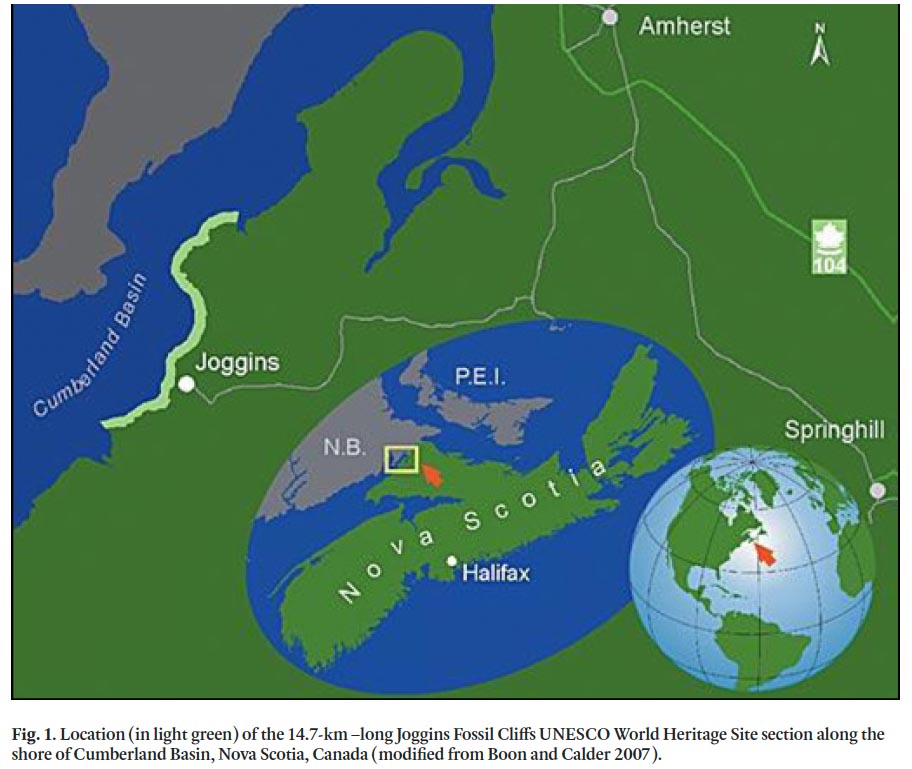
1 The Joggins Fossil Cliffs site (Nova Scotia, Canada; Fig. 1), representing the interval from about 310 to 315 Ma, was inscribed on the UNESCO World Heritage List in 2008 for its outstanding representation of Carboniferous (Pennsylvanian) biota within their environmental context. Research and discovery at the site began over 150 years ago and continues today; and with World Heritage inscription, Joggins now has an even greater presence on the global stage. Since the mid-1800s, well over one hundred scientific papers have featured Joggins, and many more mention it. Excellent accounts of the early research at Joggins, from the mid-1800s to the mid-1900s, have been undertaken by Calder (2006), Falcon-Lang (2006), Rygel and Shipley (2005), and Scott (1998). However, no in-depth review exists of recent work, from about 1950, encompassing the entire World Heritage Site, a gap that we fill with this contribution.
 Display large image of Figure 1
Display large image of Figure 1
2 Much of the research at Joggins can be placed into one of three categories: geology (e.g., sedimentology and petrology); paleobiology (including discovery and taxonomy); and paleoenvironmental reconstruction, which builds on the first two categories. We review the research within each of these categories as it relates to the World Heritage Site section and outline what we see as critical future work.
GEOLOGIC SETTING
Stratigraphy, Sedimentology and Sedimentary Environments
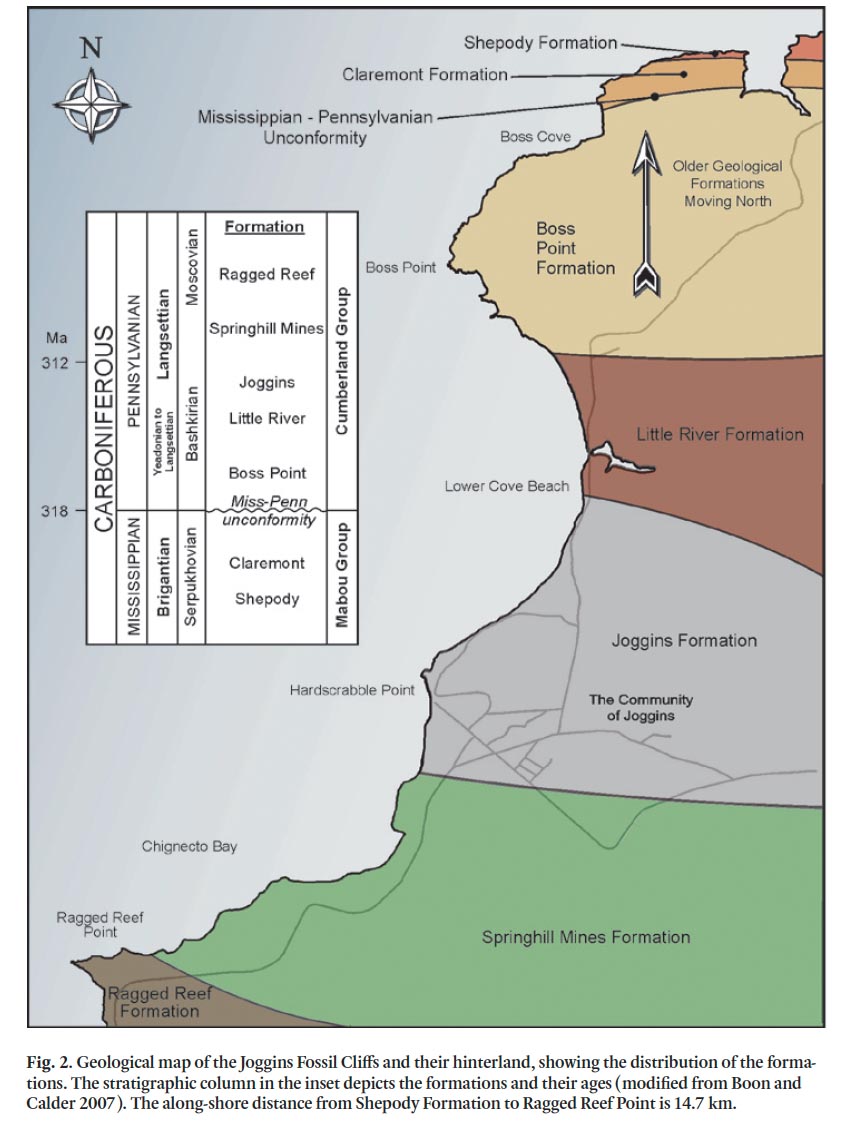
3 Some of the largest strides in research at Joggins have been in sedimentology and stratigraphy, providing a context and framework for all other recent work. The Joggins Fossil Cliffs site comprises a 14.7-km-long coastal section. The pioneering Canadian geologist, William Logan, first described the section at Joggins in 1843 and logged over 4 km of it in just five days (Rygel and Shipley 2005). Over the next 150 years, the sedimentology and stratigraphy of the section were revised by several researchers, including Dawson (1868), Bell (1912, 1914, 1944) and Copeland (1959). To date, the most complete lithostratigraphic investigations of the entire Joggins section remain those by Ryan et al. (1991) and Ryan and Boehner (1994). Ryan et al. (1991) were the first to delineate and name formations and informal members in the Cumberland Group, and this work was expanded upon by Davies et al. (2005), Calder et al. (2005a) and others. Recent tectonostratigraphic work on the oldest, Mabou Group, part of the section was the subject of a M.Sc. thesis by McLeod (2010). Today, the section is considered to include units of the Early Carboniferous (Mississippian) Mabou Group (Shepody and Claremont formations) and the Late Carboniferous (Pennsylvanian) Cumberland Group (Boss Point, Little River, Joggins, Springhill Mines, and Ragged Reef formations) (Fig. 2).
 Display large image of Figure 2
Display large image of Figure 2
4 Only sedimentary rocks are found within the section, making radiometric dating impossible; however, based on palyno-logical data from parts of the section (e.g., Dolby 1991, 2003; Utting et al. 2010) and the latest widely accepted geological timescale for the Carboniferous (Davydov et al. 2004; Davy-dov et al. 2010), the section most likely represents an interval of nearly 15 million years. Utting et al.’s (2010) detailed work is the first published study of the palynofora from the Joggins area and includes the entire World Heritage Site; their results indicate that the section ranges in age from the Brigantian (late Visean) to the Langsettian (Bashkirian?). However, Utting et al. note that palynological correlations are approximate as assemblages in the younger part of the section are of low diversity and many diagnostic taxa are absent.
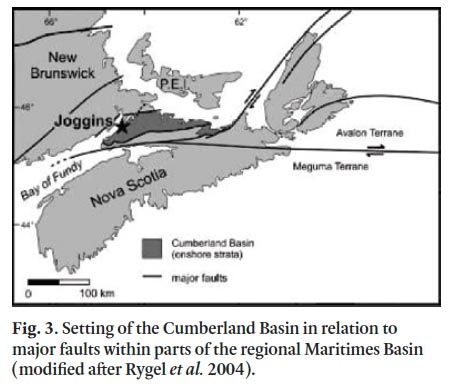
5 The sedimentary exposure at Joggins is situated in the Cumberland depocentre, which forms part of the regional Maritimes Basin complex of Atlantic Canada (Gibling 1995; Gibling et al. 2008) (Fig. 3). The strata at Joggins are tilted as part of the Athol Syncline (see Reed et al. 1993). In a seismic study of the Athol Syncline, Waldron and Rygel (2005) showed that the rapid subsidence evident in the Cumberland Basin was caused by withdrawal at depth of Mississippian evaporites, allowing for the build up and preservation of a thick sequence of sediments and the fauna and flora found within them.
 Display large image of Figure 3
Display large image of Figure 3
6 Sedimentological studies show that at sea-level highstand, this basin would have been connected, albeit distantly (based on the absence of common marine horizons), to the open-ocean as indicated by brackish incursions (Duff and Walton 1973; Archer et al. 1995; Skilliter 2001; Falcon-Lang 2005a) and drainage patterns (Gibling et al. 1992). At lowstand, the basin became restricted and was probably intracontinental (Falcon-Lang 2006). Based on paleoflow patterns in the Boss Point, Little River, Joggins, and Springhill Mines formations, paleoflow was derived variously from the Cobequid and Caledonia highlands (Rust et al. 1984; Browne and Plint 1994; Calder 1994; Calder et al. 2005a; Rygel and Gibling 2006).
7 Sedimentological studies have also revealed the changing pa leoclimatic conditions reflected in the World Heritage Site succession. Conglomeratic red beds of the Mabou Group indicate that arid conditions were widespread during the Mississippian in the Cumberland Basin (Hamblin 2001; Waldron and Rygel 2005). However, Pennsylvanian sediments mark a significant paleoclimatic change (Calder 1994). That humid conditions were common in the Pennsylvanian is reflected first in the Boss Point Formation (the basalmost Pennsylvanian formation) in part by the presence of coals (Gibling and Rygel 2008) and thick sandstone deposits from large river systems, demonstrating alternate braidplain and shallow lacustrine deposits (Browne and Plint 1994; Plint and Browne 1994). The top of the Boss Point Formation has calcareous paleosols and red floodplain deposits marking the beginning of semi-arid conditions that continue into the overlying Little River Formation (Gardiner and Gibling 2005; Gibling and Rygel 2008). Seasonal drylands were prevalent during the deposition of the Little River Formation, reflected by the lack the coal and limestone beds. In contrast, coal and limestone beds are extensive within the younger Jog-gins Formation and reflect the wetland conditions (Calder et al. 2005a). As the basin floor subsided, the lithostratigraphic succession within the Joggins Formation indicates that conditions became progressively wetter, promoting widespread growth of lycopsid trees and seed ferns that, over time, became the raw material for coal formation. While conditions were generally humid, mature paleosols and varying abundance and thickness of coal beds and red beds indicate a seasonal paleoclimate during this time (Davies and Gibling 2003).
8 The lower part of the Springhill Mines Formation records alternations between poorly drained (wetland) and well-drained (dryland) facies, similar to those of the underlying Joggins Formation (Salg and Rygel 2008). The paleosols in the Spring-hill Mines Formation were probably formed under warm and humid conditions alternating seasonally with dry conditions (Smith 1991). At the top of the section is the Ragged Reef Formation, which has been considered in less detail than the other formations (though see Way 1968 and Rust et al. 1984). However, a study by Deal (1991) indicates that paleoclimate during deposition of the Ragged Reef Formation was warm and relatively dry when compared to the generally humid paleocli-mate reflected by the older Pennsylvanian formations. Rust et al. (1984) found a lack of thick coal seams in the Ragged Reef Formation, whereas red colouration and channel sandstones became more abundant up-section within the formation. These observations were attributed to a low coeval water table and locally rapid subsidence.
9 Of particular focus in the literature has been the Joggins Formation, or "Classic Section", where nearly all of the important paleontological discoveries have been made. The Joggins Formation was deposited within an interval of less than one million years, probably during the Langsettian (Dolby 1991, 2003; Utting and Wagner 2005; Davydov et al. 2010). Davies and Gibling (2003) were first to detail the sedimentology and sequence stratigraphy of the Joggins Formation, including facies cyclicity. Davies et al. (2005) and Gibling and Rygel (2008) suggested that glacioeustasy may have been important in creating the cycles. This research was followed by numerous other sedimentological studies, discussed below, that provided a basis for the paleoenvironmental interpretation of the Joggins site, including work on facies associations, fluvial deposits, and vegetation-induced sedimentary structures (VISS).
10 Three primary facies associations have been identified within the Joggins Formation: an open-water brackish association; and two terrestrial associations, a poorly drained coastal-plain association and a well-drained alluvial-plain association (Davies and Gibling 2003). In a detailed analysis, Davies et al. (2005) described the Joggins Formation in terms of fourteen repeating cycles (or rhythms). Cycles within the 915.5-m-thick formation tend to begin with beds of coal and fossiliferous limestone, marking episodes of sea-level rise. Overlying these lithologies are sandstones and mudstones, which dominate the section and represent coastal-plain and alluvial deposits.
11 Fluvial deposits, including channel bodies, are common sedimentological features in the section, especially within the Joggins Formation. A recent study has also reported such structures in the Boss Point and Little River formations (Allen et al. 2011). Rygel and Gibling (2006) explored the numerous Jog-gins Formation channel bodies and their development in terms of intrinsic and extrinsic controls. The authors found that all the channel bodies could be defined as fixed, meandering or multistory, and that all types were represented in both coastal wetland and inland floodplain facies associations. Due to high subsidence rates (Waldron and Rygel 2005; Gibling and Rygel 2008) extrinsic factors, such as climate, eustasy and tectonics probably had a relatively small effect on the architecture and form of the channel bodies (Rygel and Gibling 2006).
12 Vegetation today has a considerable influence on the features of a landscape and, given the abundance of vegetation in the Pennsylvanian, it likely affected sediment accumulation and structures during that time also. Despite the extensive work on modern vegetation-induced sedimentary structures (VISS), one of the few studies of "fossil" VISS comes from Rygel et al. (2004). They identified seven types of VISS within the Joggins Formation: five are "simple" hydrodynamic structures that develop around a single plant or between several plants; and two types are "complex", developing from a decaying plant through infilling or soſt-sediment deformation. The VISS structures are facies-limited: hydrodynamic types are present in both dryland and wetland settings, while decay-related types only occur in the dryland settings.
Petrology
13 Joggins is in an area that was settled because of its natural resources. Early First Nations people came to fish (the name Joggins is probably derived from the Mi’kmaq word Chegoggin, loosely translated as a "place of fish weirs"; Falcon-Lang 2009); and later, Europeans settled there to mine coal (Falcon-Lang 2009; Quann et al. 2010). Scientists studying coal formation in the 1800s noted that observations at Joggins helped them resolve the mystery of how coal formed (Scott 1998). Over a century later, scientists began to quantitatively study the same coal beds in order to understand their deposition, chemical and floral/faunal composition, and hydrocarbon potential.
14 The coal beds at Joggins have been described as having been deposited on small floodplains (Kaplan et al. 1985). The coals at Joggins and those of the Maritimes Basin in general contain high levels of vitrinite and sulphur (Gibling et al. 1989; Kaplan et al. 1985; Hower et al. 2000; Skilliter 2001) and higher-than-normal heavy-metal trace-element concentrations (Kaplan et al. 1985). Kaplan et al. hypothesized that these trace elements were derived from the exposed Cobequid Massif to the south. Coal beds within the section also vary in their content of lycopsid spores versus tree-fern spores, with tree-fern spores becoming relatively more abundant over time (Hower et al. 2000). Vegetation closely reflects climatic conditions; as the paleoclimate became less humid toward the end of the Penn-sylvanian, lycopsid trees became less dominant and the tree ferns, as a group more tolerant to dryer conditions, increased in abundance (DiMichele et al. 2009 and references therein).
15 Gibling and Kalkreuth (1991) undertook petrographic analyses of Late Carboniferous carbonaceous limestones of Atlantic Canada, including material from Joggins, to determine hydrocarbon recovery potentials. While these authors found good to very good potentials for the rocks at Joggins, the beds are too thin to be economically viable (Gibling and Kalkreuth 1991). As with other studies (e.g., Archer et al. 1995; Davies et al. 2005), the authors found abundant bivalves, ostracods and vertebrate fragments within the limestones at Joggins and, based on the fauna, concluded that the units were laid down in shallow lakes or interdistributary bays with a possible marine influence (Gibling and Kalkreuth 1991). Skilliter (2001) studied coal petrology and coal geochemistry, in the process suggesting a distal marine influence for the Forty Brine section within the Joggins Formation. However, the extent of marine influence in the Joggins section has remained the subject of debate (e.g., Brand 1994; Archer et al. 1995; Skilliter 2001; Tibert and Dewey 2006) and will be further discussed below.
Future Research
16 While there have been many important advances in our understanding of the stratigraphy, sedimentology and petrology of the Joggins area, some critical gaps remain. For example, more information is required on the stratigraphy, depositional environment, and climate of the uppermost formations within the World Heritage Site, including the Springhill Mines and Ragged Reef Formations. There have also been numerous references to paleosols throughout the site, but a comprehensive study is lacking. Given the importance of these "fossil" soils to the growth of the plant life that is so exquisitely preserved at Joggins, this remains a significant gap.
PALEOBIOLOGY
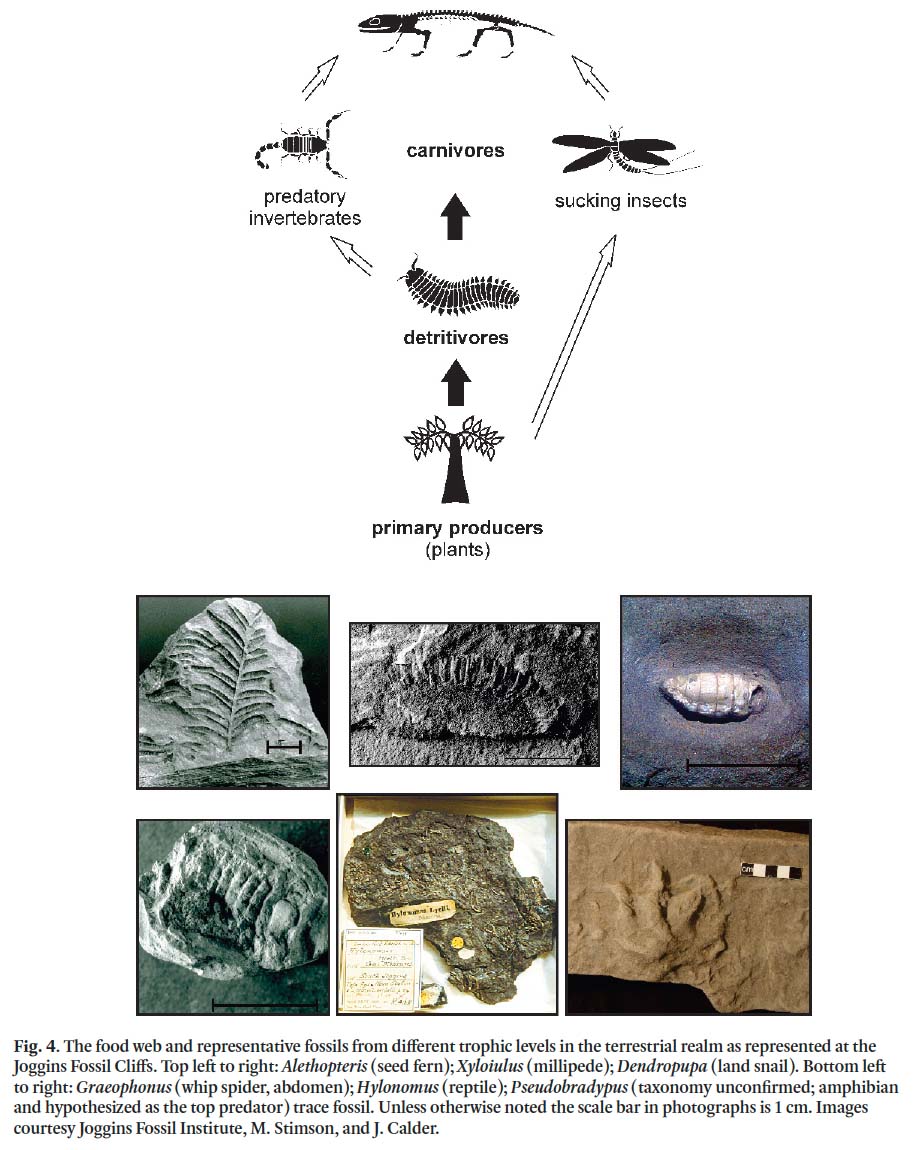
17 To date, over 200 species have been recovered from the Joggins section (Joggins Fossil Institute, unpublished data), broadly encompassing the entire food web in the terrestrial realm (Fig. 4). The assemblage represents four animal phyla, five classes of vascular plants, and three genera of protozoans (foraminifera) (tables 2.1–2.4 in Boon and Calder, 2007). Fossils from Joggins include the world’s earliest-known reptile ( Hylonomus lyelli; Fig. 4), the world’s earliest-known land snail ( Dendropupa vetusta; Fig. 4), and the tracks of the largest-known land invertebrate ( Arthropleura). The famous "fossil forests", including upright lycopsid and cordaite trees, occur throughout much of the Joggins section. These trees are of particular interest because they are preserved in situ and commonly contain remains of other organisms, collectively known as the tree-hollow fauna (see below).
 Display large image of Figure 4
Display large image of Figure 4
18 Early papers from the 1800s primarily describe new discoveries at the Joggins Fossil Cliffs. William Dawson, in particular, whose work has been summarized by Falcon-Lang and Calder (2005), Calder (2006) and Falcon-Lang (2006), extensively collected and studied fossils from Joggins. Many of the more recent discoveries and taxonomic studies are of the vertebrates, mainly amphibians, including the extinct groups of the microsaurs and labyrinthodonts; (e.g., Carroll 1963, 1966, 1967; Stevenson and Stevenson 1966; Stevenson 1967; Baird 1982; Godfrey et al. 1987, 1991; Milner 1996; Reisz and Modesto 1996; Holmes et al. 1998; Robinson et. al 2005; Holmes and Carroll 2010). Fewer papers have focussed on vertebrate and invertebrate trackways, but include those by: Ferguson (1966, 1975) on Arthropleura trackways ( Diplichnites); Sarjeant and Mossman (1978), who reviewed vertebrate trackways; and Mossman and Grantham (1996) on an amphibian trackway ( Dromillopus). Dawson (1890) and Matthew (1903, 1905) outlined 13 "species" of tetrapod trackways, but there has been very little recent work synthesizing the knowledge of vertebrate and especially invertebrate traces. It should be noted that Lucas et al. (2005) and Stimson and MacRae (2010) produced reviews of tetrapod and Diplichnites and Kouphichnium trackways, respectively, but these remain in abstract form. Dafoe et al. (2011) have also reported on the invertebrate traces at Joggins.
19 Very few studies have considered microfossils from Jog-gins, with the exception of palynological studies. Aside from broadening the evidence for prevailing flora, spores and pollen have provided a principal criterion for dating the Joggins Formation (e.g., Dolby 1991, 2003; Utting et al. 2005; Utting et al. 2010). Other micropaleontological work has involved foraminifera and ostracods; for example, Tibert and Dewey (2006) focused on the taxonomy of the Joggins microfossils, specifically the ostracods, which are especially abundant within the limestones. They assigned most of the ostracods they found in the Joggins Formation to a new genus, Velatomorpha, which they considered to have lived in brackish, estuarine-type paleoenvironments. This conclusion was based on faunal associations of Velatomorpha with other taxa, including ostracods they attributed to Carbonita spp. Carbonita occurs in a range of non-marine to brackish conditions, supporting the conclusion of Archer et al. (1995) that deposition of at least some of the aquatic beds took place in brackish water.
20 There has also been relatively little work on the taxonomy of Joggins plant material. Falcon-Lang (2005b) described ad-pressed tree-fern trunks and Mosle et al. (2002) described seed-plant cuticles, but broader studies have yet to be published. A global review of the Coal Age macroflora has been undertaken by R. Wagner (Palaeobotanical Museum (IMGEMA-Jardín Botánico de Córdoba, Spain) and remains unpublished.
 Display large image of Figure 5
Display large image of Figure 5
Future Research
21 The rates of publication and taxonomic discovery at Joggins have been explored by Falcon-Lang (2006), whose analysis indicates that the saturation point of discovery has not yet been reached. The potential for finding new species is still relatively high, making Joggins an exciting place for paleontologists. With an increased global awareness of the Joggins site since its inscription on the World Heritage List in 2008 and the presence of the Joggins Fossil Institute (which houses an interpretive gallery, fossil collection, and hosts guided tours of the site), discovery of new specimens has already increased from Falcon-Lang’s (2006) count of about 148 species to about 200 species. Some of this increase can be accounted for through recent taxonomic revision of previously described taxa, but this number also represents a true increase of new material.
22 Taxonomy remains the basis of almost all paleontological research. Future taxonomic research will be especially critical in several areas. Many reviews and revisions for the tetrapod fauna exist (e.g., Carroll 1964, 1969, 1970a, 1970b, 1982, 1992, 2001; Milner 1987), but as noted, relatively little work has been published on the floral and trace-fossil taxonomy, and thus these two areas represent significant current gaps.
23 From the authors’ perspective, the next major steps for research at the site should be in the field of evolution, of which taxonomic work and morphological analysis of traits comprise the foundation. Hylonomus lyelli represents an important evolutionary innovation in the history of vertebrates, reflecting their full transition from water onto land; these first amniotic tetrapods have been the subject of numerous papers (e.g., Carroll 1992; Milner 1996; Reisz 1997; Benton and Donoghue 2007). One recent study has linked tetrapod diversification during the Late Carboniferous to rainforest collapse in the former Euramerican part of Pangea (Sahney et al. 2010); micro-evolutionary studies of the various taxonomic groups within the well-preserved Joggins assemblages, representing an approximately 15-million-year interval, could add new insights into such research. In order to build capacity for these kinds of studies, a critical focus must be on collecting and repatriating stratigraphically constrained and well-preserved fossils. (Note that, as of 1982, all fossils found in Nova Scotia legally belong to the province and a Heritage Research Permit must be obtained in order to collect specimens. For permit information see www. museum.gov.ns.ca/fossil/protect/permits/html. Aſter study, these specimens must be returned to the province and will be added to the collection of the Nova Scotia Museum. The Jog-gins Fossil Centre houses a collection of Joggins fossil material that is on renewable loan from the Nova Scotia Museum and is currently the primary repository for all new Joggins material).
24 Linkages between oxygen and body size have been tested in numerous studies, most notably by Chapelle and Peck (1999) and Spicer and Gaston (1999), suggesting that gigantism (at least in amphipod crustaceans) is correlated with the concentration and partial pressure of oxygen. However, debate continues as to whether oxygen is a covariant and whether other environmental factors such as temperature are involved (Spicer and Gaston 1999). In insects, maximum body size is controlled by tracheal diffusion; therefore an increase in oxygen partial pressure would increase metabolism and allow for a larger body size (Graham et al. 1995). This factor most likely explains gigantism among insects in the Carboniferous and the ensuing extinction of such forms in the Permian when the partial pressure of oxygen fell to approximately two-thirds the value today (Spicer and Gaston 1999). We see the invertebrate fauna at Joggins as a potential testing ground for tracking linkages between atmospheric oxygen concentration, gigantism, and perhaps even diversity.
PALEOECOLOGY
25 An important factor contributing to the designation of the Joggins Fossil Cliffs as a World Heritage Site was that fossils found there are preserved in situ that is, in their paleoenvironmental context. Both sedimentological and faunal analyses serve as a foundation to reconstructing paleoenvironments. In general, the Late Carboniferous world evokes images of humid, rainforest-type settings (Fig. 5); this representation, however, is based on "classic" Carboniferous localities in eastern North America and western Europe that were situated in the paleotropics. We now know that the climate during this time period was more complicated, with some areas globally subhumid to perhaps even semi-arid (DiMichele et al. 2010). Geophysical data suggest that Joggins was near the paleoequator during the Carboniferous (DiVenere and Opdyke 1991) and therefore falls into the "ever-wet tropical" climatic belt (DiMichele et al. 2010) or "tropical peat forest biome" (Fig. 6).
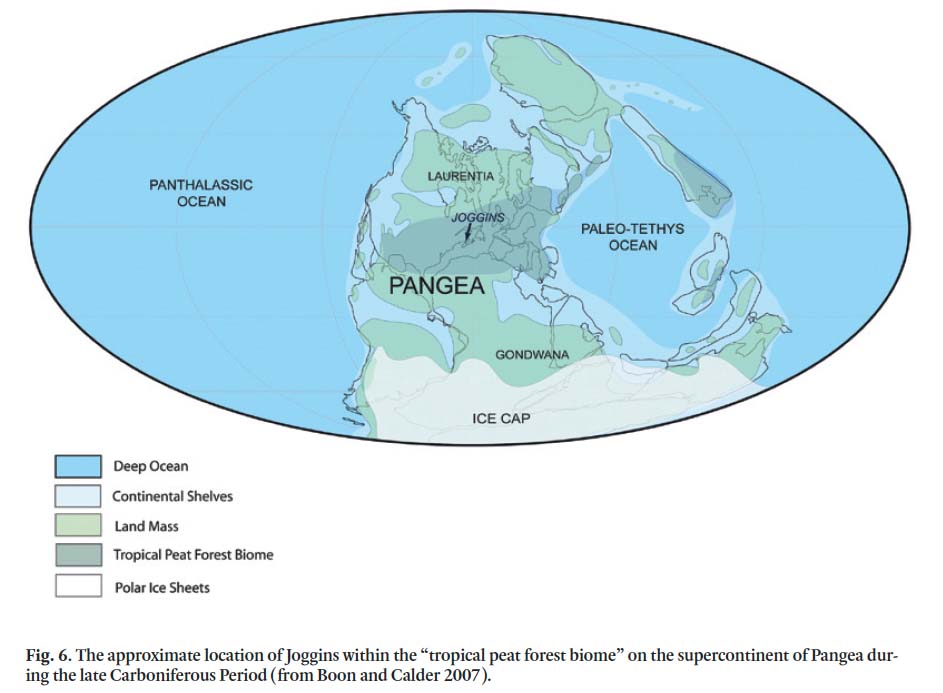 Display large image of Figure 6
Display large image of Figure 6
26 As outlined above, sedimentological studies have revealed climatic changes throughout the World Heritage Site. In particular, the well-studied Joggins Formation shows cyclicity of facies associations and thus of paleoenvironments. Falcon-Lang (1999) and Falcon-Lang et al. (2004, 2006) related the sediments and their contained sedimentary structures and fossil assemblages to specific paleoenvironments. The Joggins Formation biota exhibits a cyclic succession that was probably climatically driven by glacial-interglacial cycles (Falcon-Lang 2003a). The succession has three primary components: a cordaite dry-land community, a lycopsid wetland rainforest community, and an aquatic paleoenvironment representing perhaps proximal influence of maximum transgressions (Falcon-Lang 2003a; Falcon-Lang et al. 2006). Many comprehensive studies exist on the terrestrial realm, but fewer have focussed on the aquatic setting, and most detailed studies of paleoenvironment have been primarily within the Joggins Formation. Details of the three paleoenvironment types follow.
Terrestrial Drylands
27 The red beds of the Joggins Formation and the older Little River Formation are evidence that wetlands were not the sole environment-type exhibited within the World Heritage Site. Terrestrial drylands are associated with well-drained alluvial plain facies described by Davies et al. (2005) and were dominated by dryland plant communities composed primarily of pteridosperms (seed ferns) and large cordaitalean trees (Falcon-Lang and Scott 2000; Falcon-Lang 2003b; Falcon-Lang 2007). The trees belong to an extinct order of gymnosperms, the Cordaitales, related most closely to conifers; however, chemosystematic and microstructural investigations of cordaites, other conifers, and seed ferns revealed that cordaites were no more closely related to conifers than they were to seed ferns (Mosle et al. 2002).
28 Cordaitaleans reached heights of approximately 45 m; grew under humid tropical conditions; lived in an environment where wildfire occurred; and probably lived primarily in an upland setting (Falcon-Lang and Scott 2000; Falcon-Lang and Bashforth 2004), an important finding as this indicates that upland floras may have existed earlier than previously thought (Falcon-Lang and Scott 2000). Cordaitalean trees of the Boss Point Formation have also been noted as potentially the earliest evidence of log jams in the fossil record (Gibling et al. 2010), but the jams have only been noted (Calder 1998; Falcon-Lang and Scott 2000; Gibling et al. 2010) and have yet to be documented in detail.
29 Very rare cuticles of indeterminate scorpions and eurypterids have been found in the dryland facies of the Joggins section (Stankiewicz et al. 1998; Falcon-Lang et al. 2006). Interestingly, these cuticles were used by Stankiewicz et al. (1998) to help determine the contribution of animal remains to the formation of kerogen, which may be higher than previously thought.
30 In a quantitative analysis of fossils from the Joggins Formation using species diversity indices, Falcon-Lang (2003c) showed that growing conditions in the terrestrial drylands were stressed and that community composition differs considerably from wetland communities within the same formation. That author noted that sequence stratigraphy of the Joggins Formation suggests that the dryland flora grew in continental-interior environments (lowstand aridity) as opposed to wetland flora that grew in humid coastal settings (highstand humidity). A fossil assemblage from a 2-m-thick sandstone bed within the Joggins Formation displays a unique dryland assemblage of land snails ( Dendropupa), archanodontid bivalves, and tetrapods such as Baphetes, a fauna probably representing a dry-land "waterhole" setting, the first of its kind to be described in the Carboniferous (Hebert and Calder 2004; Falcon-Lang et al. 2004). Falcon-Lang et al. (2004) considered the seasonal drainages and waterholes of present-day central and northern Australia to be modern analogues of this paleoenvironment.
Terrestrial Wetlands
31 The wetland settings within the Joggins section, in which the standing lycopsid trees (specifically, Sigillaria and Lepidodendron) are primarily preserved, are interpreted as representing a disturbance-prone system with heavy rainfall and seasonal flooding (Falcon-Lang 1999, Calder et al. 2006). Wetlands are associated with the poorly drained coastal plain unit of Davies et al. (2005), where communities were composed of amphibians, reptiles, arthropods, pteridosperms, ferns, lycopsids, cordaita-leans, and calamiteans (Falcon-Lang et al. 2006). Secondary succession is apparent within the wetland environment: as lycopsids periodically drowned by flooding events, Calamites (a disturbance-tolerant relative of the modern day horsetail Equisetum) would regenerate and flourish (Calder et al. 2006).
32 Cordaitalean trees are a primary feature of dryland and upland communities, but they were also found in wetlands (Falcon-Lang 2005a). It was therefore once thought that cordaitalean trees may have represented early mangrove-type communities (Raymond and Phillips 1983). Falcon-Lang (2005a) assessed this hypothesis and found that, while the trees could grow in close proximity to brackish seas and sedimentological evidence suggests that at times they were partially submerged, no evidence to date suggests that they were mangroves in the strict sense.
Coastal Plains and Seas
33 The aquatic environment and its degree of marine influence has been the subject of continued debate for decades. Until recently, the aquatic fauna of the Joggins Formation could not provide further information regarding the paleoenvironment because no diagnostic marine (salinity ≈ 35 ppt) or freshwater (salinity < 0.5 ppt) organisms had been identified. The majority of the fauna includes: bivalves with a freshwater affinity ( Naiadites, Curvirimula, Archanodon); poorly preserved agglutinated estuarine (≈ marginal marine; salinity ≈ 0.5–30 ppt) foraminifera; ostracods ( Velatomorpha) with an estuarine affinity; scales and bone material (teeth, ribs, etc.) of bony (rayfinned and lobe finned) and cartilaginous fish; microconchid worm tubes (long misidentified as Spirorbis polychaete worms: Taylor and Vinn, 2006); shrimp ( Pygocephalus); and horseshoe crab trackways ( Kouphichnium).
34 Previous studies of the calcareous shales in the Joggins Formation suggested a marginal marine (≈ estuarine) setting based on: faunal composition (see above); association of carbonaceous strata with rooted mudstones and coals; and alluvial-channel deposits (e.g., Duff and Walton 1973; Gibling and Kalkreuth 1991). A marginal marine setting also is supported by two later studies that focused on faunal and sedimentological analyses of different beds within the Joggins Formation. Trace fossils (including Kouphichnium and annelid traces belonging to the genera Cochlichnus and Treptichnus) and agglutinated foraminifera, recovered from sandstone and mudrock beds in the middle of the Joggins Formation, suggest the presence of marine-influenced waters in an estuarine-type setting (Archer et al. 1995). Sedimentological structures such as ladder-back and double-crested ripples found in the beds are also indicative of, but not limited to, a tidally influenced setting (Archer et al. 1995). A second study supporting this conclusion was based on faunal associations and morphological features of the ostracods found within limestone beds of the Joggins Formation (Tibert and Dewey 2006).
35 The only geochemical analysis of fossil material from the Joggins section was on Naiadites bivalves from the upper part of the section (Brand 1994). The strontium, magnesium, sodium, manganese and iron contents significantly differed from those of coeval tropical marine molluscs, suggesting that the Joggins material is from a near-shore lacustrine (non-marine) setting with slightly dysaerobic waters (Brand 1994). This finding is in direct contrast to the previous work mentioned and thus the degree of marine influence at the site remained a matter for debate until recently. Grey et al. (2011) studied the limestones throughout the entire Joggins Formation and found evidence to suggest a waning marine influence. At the bottom of the Formation, echinoderms, brachiopod fragments, and pyritized ostracods were found in thin section, indicative of marine conditions; closer to the top of the Formation there were no echinoderms and many more non-marine bivalves ( Naiadites and Curvirimula) were present (Grey et al. 2011).
The "Tree Hollow Fauna"
36 Common features of the Late Carboniferous tropical landscape evidenced by the Joggins succession included wildfire (perhaps promoted by high atmospheric-oxygen content) and seasonal and/or storm flooding events. Both features play a role in possible explanations for the famous "tree hollow fauna" of Joggins. Studies of fusain (fossil charcoal) in the Joggins section indicate that wildfires were frequent (Falcon-Lang 1999, 2000; Hower et al. 2000). There were two types of fire-prone plant communities: cordaite/conifer forests occurring in dry, upland niches; and a wetland community dominated by medullosan pteridosperms (seed ferns) and tree-sized club mosses (lycopsids), such as Sigillaria and Lepidodendron (Falcon-Lang 1999). Flooding is evidenced to some extent by upright sediment-filled trees whose trunks contain some of the most famous discoveries at Joggins, including Hylonomus lyelli.
37 Animals such as Hylonomus and at least eleven other tetrapod taxa, as well as detritovorous invertebrates and insects, may have been killed by wildfires (Falcon-Lang 1999) or flooding events that entombed the fauna in the trees, where at least some of the tetrapods may have been living/denning (Calder et al. 2005b). Fragments of plant material are also frequently found within sediment-filled trees that may have been drowned by large-scale, perhaps seasonal, flooding events (Calder et al. 2006). Lycopsids, the dominant tree of the wetland community, would rot from the inside and create a hollow trunk that may have been used as a living space for some animals (Calder et al. 2005b). Further flooding events would fill the hollowed tree with sediment and plant material, entombing the animals where they denned (Calder et al. 2005b).
38 Denning is one of three primary scenarios used to explain the occurrence of the tree hollow fauna. Two other scenarios have been proposed. In one, the animals (and plants) are postulated to have been washed into the trunk aſter death by flooding. In the other, the animals are envisaged as having fallen into partially buried hollowed trunks; this is known as the "pitfall theory" (Calder et al. 2005b). Early views, especially of Lyell, favoured the pitfall explanation, whereas a more recent study has favoured the denning scenario (Calder et al. 2005b); but the taphonomic and sedimentological details of the latter have yet to be published.
Future Research
39 In terms of improving the understanding of the paleoenvironmental context of the Joggins section (i.e., the entire World Heritage site), we see three areas as most promising. First, a need clearly exists to expand the research outside of the Jog-gins Formation; for instance, the log jams at Boss Point certainly warrant further study. Second, geochemical, specifically isotopic and trace-element studies, may be particularly useful in tracking potential changes in paleoclimate throughout the time represented by the World Heritage Site section. Such work would also provide an opportunity to explore the correlation of climatic and paleoenvironmental conditions with evolutionary patterns and biodiversity. Geochemical studies, however, must proceed with caution as diagenetic alterations can severely affect results. As Falcon-Lang (2003a) indicated, considerably more research is required to understand the impact of medium-term global environmental change on biodiversity, a subject of particular relevance today as the world is in the midst of a biodiversity crisis (e.g., Lawton and May 1995; Pimm et al. 1995; Thomas et al. 2004; Barnosky et al. 2011). Finally, publication of sedimentological and paleontological evidence would be useful in moving forward with hypotheses on the intriguing and well-preserved tree hollow fauna.
40 The Carboniferous Period, which has supplied the world with its main source of energy over the past few centuries, witnessed the highest atmospheric oxygen concentration in Earth history; and was a key time in evolution as vertebrates in the form of reptiles became independent of the aquatic environment. Few, if any, broadly based comparative analyses or synthesis of Carboniferous sites around the world exist. A couple of publications deal with specific aspects. One of these, by Carroll (1994), dealt with the tetrapods and noted that the East Kirkton (Scotland) tetrapod faunas were most similar to those of Joggins. The other, by DiMichele and Falcon-Lang (2011), reviewed aspects of Carboniferous to earliest Permian fossil forests preserved in growth position. In situ forests preserved in a stratigraphic series as at Joggins can, with limitations, provide unique knowledge of forest ecology and spatial patterns within specific habitats.
41 Perhaps the best example of a synthesis of global Carbon-iferous sites is an unpublished analysis written as part of the Joggins Fossil Cliffs nomination package to UNESCO for World Heritage status (Falcon-Lang 2002). This analysis compared other globally significant Pennsylvanian sites to Joggins. While extremely useful, the report focused on the prerequisites and benchmarks derived from the Questions and Recommendations of the International Union for Conservation of Nature (IUCN) for evaluation of fossil sites. Overall, nine sites were considered under three primary categories: the fossil record of biodiversity; the nature, quality and variability of the fossil archive; and the permanence (integrity) and scientific impact of the site. It may now be useful to expand that review from these three categories to include a synthesis on what we know about the Late Carboniferous world in general – from climate to paleogeography to faunal changes – based on critical sites globally.
CONCLUSION
42 Much of the recent research related to the Joggins Fossil Cliffs falls into three main categories: general geology (stratigraphy and sedimentology); paleobiology (taxonomic description/ discovery); and paleoecology. This research helped to form the foundation and breadth of knowledge that allowed Joggins to be recognized as a UNESCO World Heritage Site. Within this framework we have highlighted some gaps and opportunities that include, but are certainly not limited to, studies on: the uppermost formations (Springhill Mines and Ragged Reef) within the World Heritage Site; paleosols; floral and trace fossil taxonomy; and microevolutionary patterns exhibited within the Site, with potential to couple this with geochemical work. We also recognize that many more areas of potential research exist.
43 Falcon-Lang et al. (2006) highlight that publication records have dramatically increased over time since the 1850s and projected that trend to continue throughout the past decade. We can now confirm that their predictions were indeed correct and were in fact exceeded, with over 40 publications on the Joggins site and/or material in the past decade (2000–2010). With the chance of discovery remaining high and the opportunity to use the site as a benchmark of the Carboniferous world, the Jog-gins locality will remain an area of interest to researchers for a long time to come.
This work was funded by a National Sciences and Engineering Research Council PDF to Melissa Grey and a National Sciences and Engineering Research Council Discovery Grant to Zoe V. Finkel. Jenna Boon (JFI) and two anonymous reviewers provided constructive feedback. We especially thank Co-Editor Rob Fensome and Associate Editor Martin Gibling for their insightful comments and suggestions which have significantly improved this paper.