Articles
Hypothesized Cambrian medusae from Saint John, New Brunswick, reinterpreted as sedimentary structures
ABSTRACT
More than a hundred radial and discoidal structures occur on bed tops of shales and very fine sandstones of the Cambrian (Series 3) King Square Formation in New Brunswick, Canada. These structures typically contain a central sediment plug, radial lineations that extend outward from the central plug, concentric rings, and a broad trough surrounding or underlying ring margins. Originally interpreted as fossils of scyphozoan medusae, these structures could represent one of only a half-dozen mass strandings documented from the fossil record. Instead, re-evaluation of their sedimentology and morphology suggests that they are likely sedimentary structures known as Astropolithon. These sand-volcano-like structures formed by subsurface blistering, cracking, and failure of a near-surface or surface bed, triggered by the upward movement of gases or other fluids from underlying beds.
RÉSUMÉ
Plus d’une centaine de structures radiales et discoïdes apparaissent au sommet des couches de schiste et de grès très fins de la formation King Square du Cambrien (Série 3), du Nouveau-Brunswick, au Canada. Ces structures contiennent généralement un culot sédimentaire central, des linéaments radiaux qui se déploient vers l’extérieur à partir du culot central, des anneaux concentriques, et une vaste cuvette qui ceinture le bord des anneaux ou qui se trouve en dessous. On estimait initialement qu’il s’agissait de méduses scyphozoaires, mais ces structures pourraient correspondre à l’une de la demi-douzaine d’échouages massifs de l’ichnofaune dont les vestiges fossiles ont été retrouvés. La réévaluation de leur contexte de sédimentation et de leur morphologie porte plutôt à croire que ce sont vraisemblablement des structures sédimentaires connues sous le nom d’astropolithon. Ces structures apparentées à un volcan de sable ont été formées par l’apparition d’alvéoles souterraines, le fendillement et la rupture d’une couche près de la surface ou à la surface, et qu’aurait provoquée le déplacement vers la surface de gaz ou d’autres fluides provenant des couches inférieures.
[Traduit par la redaction]
INTRODUCTION
1 Cnidarian medusae are important components of pelagic ecosystems, but are among the rarest of fossils — partly because most of them are completely soſt-bodied, and partly because once buried or stranded they are attractive resources for scavenging fauna and microorganisms. Thus, when preserved they not only signal that exceptional preservation has occurred, but also the opportunity to evaluate a broader spectrum of the biota and its paleoecology at a given moment and place in time. For example, throughout earth history, genuine scyphozoan, hydrozoan, or cubozoan medusae are only known from ≤10 deposits, all of which are significant or potential Konservatlagerstätten (Young and Hagadorn 2010). Deposits containing more sclerotized medusae such as porpitids (chondrophorines) and conularids are at least 1–2 orders of magnitude more common than their soſter counterparts (Fryer and Stanley 2004; van Iten et al. 2006).
2 Yet there are abundant reports of isolated or rare medusa-like structures, or of problematic fossils attributed to medusae (see reviews in Häntzschel 1975; Young and Hagadorn 2010). Most of these structures have a circular shape and radial or concentric ornamentation, and may have internal structures that resemble the internal cavities or organs of medusae. Whereas some of these reported discoids may represent genuine fossil discoveries, most represent other phenomena. The majority are inorganic sedimentary structures (e.g., Plummer 1980; Jensen et al. 2002), trace fossils (e.g., Häntzschel 1970; Fürsich and Kennedy 1975; Seilacher 1984), or other organisms (e.g., Conway Morris and Robison 1988; Dzik 1991; Landing and Narbonne 1992).
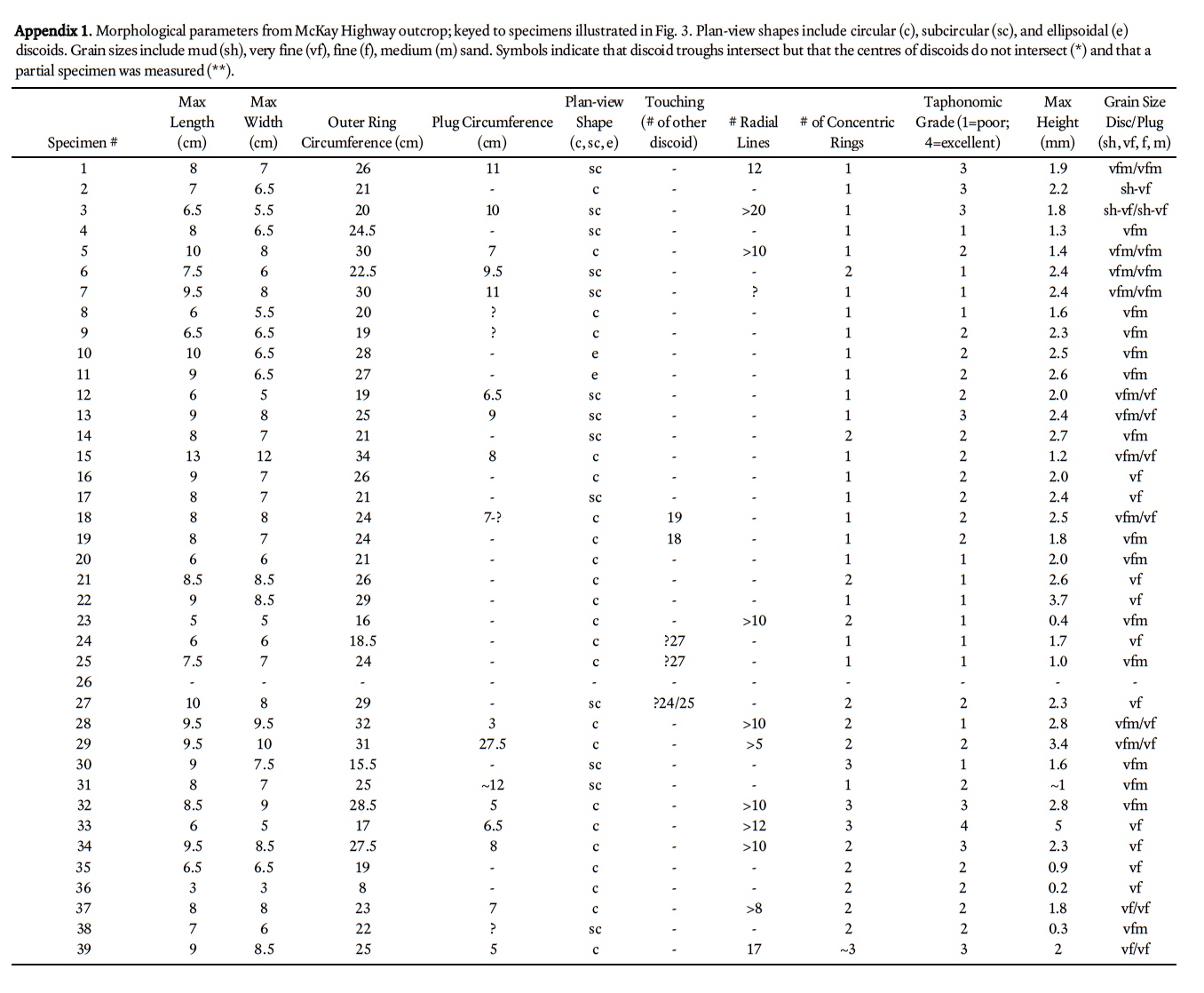
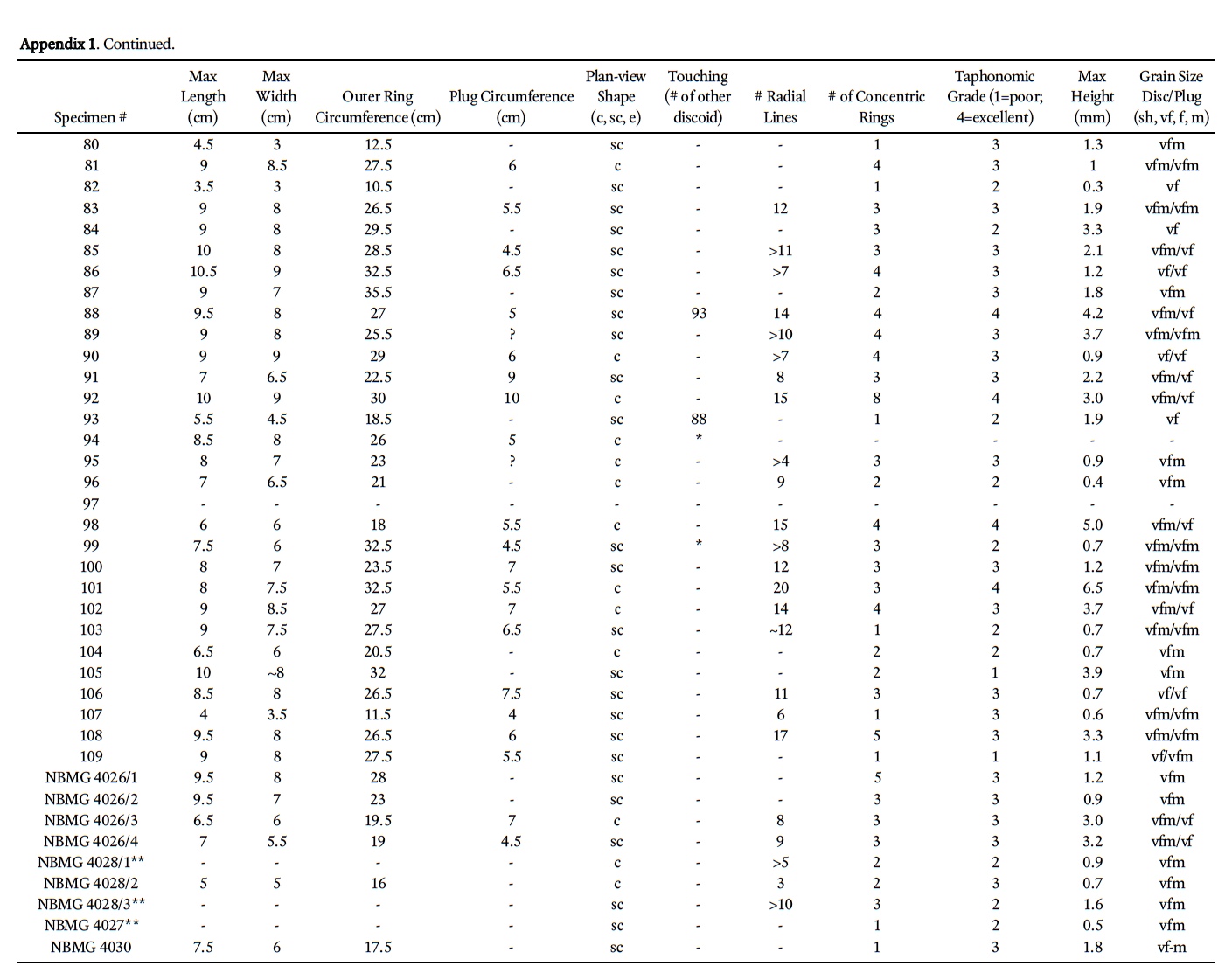
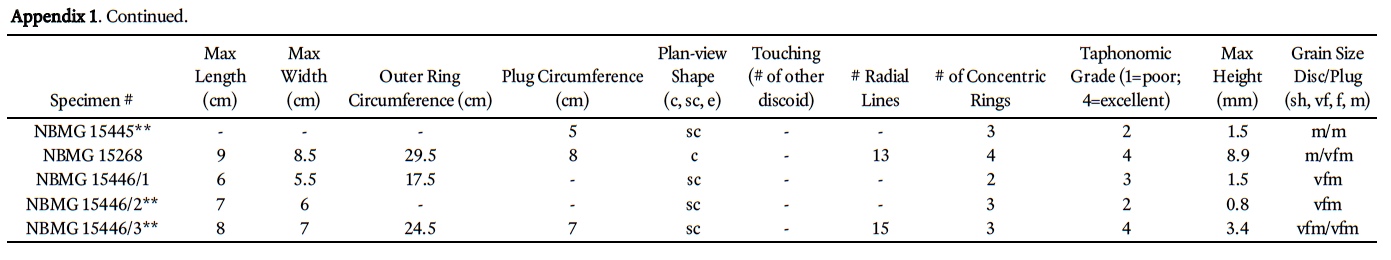
3 The Cambrian is the preservational acme of genuine medusae-bearing deposits, with at least four Middle Cambrian deposits that bear abundant or well-preserved medusae (Devereux 2001; Hagadorn et al. 2002; Cartwright et al. 2007, Hagadorn and Belt 2008; Lacelle et al. 2008). A fiſth possible deposit exists in coeval strata of southern New Brunswick, in which more than a hundred discoidal to radial structures occur; these were informally described and hypothesized to represent composite molds of scyphozoan medusae (Pickerill 1982, 1990). Since this early work, new medusae-rich deposits have been discovered and described (e.g., Young et al. 2007), and additional investigations of discoidal sedimentary structures (McIlroy and Walter 1997; MacGabhann et al. 2007) and discoidal-domal microbial structure formation have been published (York et al. 2005; Dornbos et al. 2007). On the basis of this later published work and in the context of associated sedimentologic, taphonomic, and ichnologic information, we formally describe and reinterpret the New Brunswick hypothesized medusae.
GEOLOGICAL CONTEXT
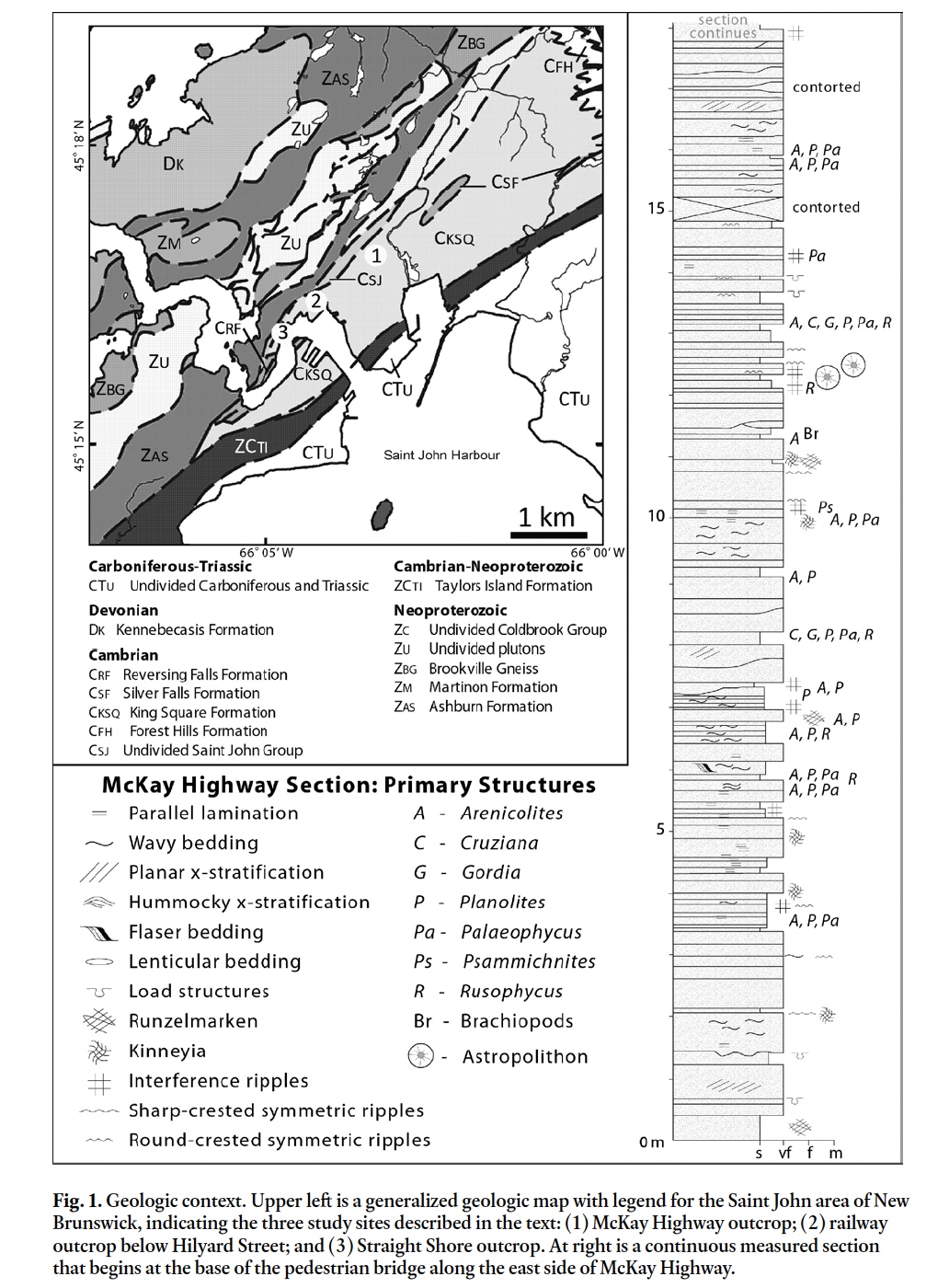
4 Disc-like structures previously considered to be medusae occur in outcrops of the Cambrian (Series 3; Furongian) King Square Formation in Saint John, New Brunswick (Fig. 1). The main study outcrop is along McKay Highway at 45° 17.09N, 66° 03.32W (Fig. 1; site 1). Broadly similar, but unrelated discoidal trace fossils (Fig. 2) occur at other sites in the King Square Formation (sites 2-3 in Fig. 1), and have also evoked comparison to medusae, for example by Matthew (1891) as Medusichnites. The latter were originally hypothesized to be traces made by medusa, but are more likely algal tool-marks or other trace fossils (Fillion and Pickerill 1990). Although alternative strati-graphic nomenclature has been proposed for the King Square Formation (Landing 1996, 2004), we use the original formation names here to foster comparison of our work with the earlier work on this unit.
5 The studied outcrops expose the middle part of the King Square Formation and overlie strata of the Forest Hills Formation bearing Eccaparadoxides eteminicus (Pickerill 1990; Kim et al. 2002) and underlie strata bearing Agnostus pisiformis trilobites (Hayes and Howell 1937). The outcrops also contain undescribed pre-Agnostus pisiformis acritarch assemblages that confirm the timing of their deposition to the Cambrian Epoch 3 (Palacios et al. 2009). In the Saint John area, the King Square Formation is dominantly fine-grained quartz arenite, interbedded with minor siltstone and shale. Many bed tops are characterized by interference ripples; beds are dominated by horizontal to wavy lamination, commonly capped by wavy lamination, unidirectional cross-lamination or climbing ripple cross-lamination (figs. 4, 6 in Tanoli and Pickerill 1989). Bed-forms typical of tidally influenced settings, such as flaser and lenticular bedding, are also common. Locally, load structures and ball-and-pillow structures are abundant. In the study area, arenites also contain linguloid brachiopods and/or are mantled by rare shallow-tier trace fossils such as Palaeophycus, Planolites, Psammichnites and Rusophycus; shales are highly bioturbated and have a diverse array of traces at shale/sandstone bed interfaces (Fig. 1; Pickerill 1982, 1990; Pickerill and Blissett 1999). Studied exposures are hypothesized to represent deposition in shallow subtidal inner- to mid-shelf environments above storm wave base (Tanoli and Pickerill 1989). Our observations of primary structures and bedding at the McKay Highway outcrop are internally consistent with this interpretation.
6 In the field, bedding planes of strata that contain the discoidal structures were cleaned with bleach, gridded, mapped, and measured. Several samples were collected for serial sectioning and for repositing in the New Brunswick Museum (NBMG). Where it was not possible to collect part-counterpart specimens, discoidal structures were cast with silicone and these were reposited. Specimens reposited at the New Brunswick Museum include two silicone molds of structures preserved on bed tops (NBMG 4026/4-6), one bed sole and corresponding latex mold (NBMG 15268/1-2), and two bed top and one bed sole slab(s) in which the discoidal structures have been sectioned (NBMG 4026/1-3; NBMG 4027).
OBSERVATIONS
7 Discoidal structures are preserved on the bed tops of two horizons in convex epirelief, and more rarely as casts on bed soles in concave hyporelief. Discoid-bearing beds are dominantly composed of thinly interbedded to laminated, mica-ceous, very fine sand and shale. Runzelmarken and Kinneyia, two structures that can be produced by microbial or inorganic processes (Hagadorn and Bottjer 1997; Porada et al. 2008) are common at the McKay Highway section (Fig. 1).
 Display large image of Figure 1
Display large image of Figure 1
8 The lower (main) discoid-bearing bed is about 10 cm thick and bears Gordia, Palaeophycus, and ?Rusophycus [bedding plane bioturbation index (bbi) of 1; ichnofabric index (ii) of 0], as well as ~10 cm wavelength, 1.5 cm high interference ripples and wavy to straight-crested oscillation ripples (Fig. 3). The upper discoid-bearing bed varies from 1–8 cm thick and bears Arenicolites, Palaeophycus, Planolites, and Rusophycus (bbi: 3, ii: 1), as well as ~8 cm wavelength, 1.5 cm high, straight- to sinuous-crested, mud-draped oscillation ripples. Discoids occur on both ripple crests and troughs of each of these beds.
 Display large image of Figure 2
Display large image of Figure 2
 Display large image of Figure 3
Display large image of Figure 3
9 In the field, 139 discoidal structures were examined, as were 14 New Brunswick Museum specimens (Fig. 4). Among all of this material, only five examples occur of two discoids touching one another, and no examples occur of more than two discoids touching one another. Where two discoids abut one another, the edges of their rings and outer trough (where present) always coalesce (Fig. 4j–k), rather than overlap.
10 Discoids exhibit a range of preservational quality on the outcrop (Appendix 1). To help constrain our interpretations to only the best-preserved specimens, we ranked all discoids according to the following taphonomic scheme: (1) recognizable, but poorly preserved specimens (14%); (2) specimens in which 20–60% of the disc area is exfoliated, fractured, or missing (34%); (3) specimens with less than 20% of the disc area exfoliated, fractured, or with missing pieces (43%); and (4) "complete" preservation (9%).
11 Among the better preserved specimens (categories 3 and 4 above), discoidal structures commonly contain four distinctive features, including a central sediment plug, radial lineations that extend outward from the central plug, concentric rings, and a broad trough surrounding (or rarely, underlying) ring margins (Fig. 4a-l). These features co-occur in four ways, as seen in plan view; (i) as a convex or flat-topped ring or as concentric rings; (ii) as concentric rings with radial lineations and a circular sediment plug; (iii) as concentric rings with radial lineations, a circular sediment plug and a surrounding trough; or (iv) as concentric rings with radial lineations and a trough (i.e., no sediment plug visible).
12 Discoids have a mean diameter of 7.4 cm, but vary from circular (51%; Fig. 4j, m) to subcircular (47%; Fig 4h, k) to ellipsoidal (2%) in plan view. They may be as narrow as 2.5 cm or as wide as 13 cm. The mean relief of discoids is 2.1 cm, with some as low as 0.2 cm and one specimen exhibiting 8.9 cm of relief above the bedding plane. The discs are inconsistent in shape and size, with some well-preserved specimens only exhibiting a half-circle of a disc (e.g., see "half-discoid" in Fig. 4i).
13 Specimens that bear concentric rings typically have 2–3 rings (e.g., Fig. 4c), but others may have no rings at all, and some may have as many as eight rings (e.g., Fig. 4l). Those specimens that bear radial structures may have as few as 2 or as many as 20 such structures; the average is 10. Radial structures (Fig. 4a–d, i, j, l) always cross-cut concentric rings, and in many ring-bearing specimens extend beyond the outermost concentric ring (Fig. 4a, c). Radial structures in some specimens extend at least 1 cm below the bed surface. Radial structures also appear to be v-shaped in cross-section, are filled with sediment, and inconsistently cross-cut parts of the discoid; for example, in some specimens the radial structures cross-cut inner concentric rings 1–3, but other radial structures cross cut both these inner rings and some outer rings (e.g., Fig. 4c, l).
 Display large image of Figure 4
Display large image of Figure 4
14 Troughs surrounding discoids may intersect (see Fig. 3, specimens 94 and 95, and Fig. 4j, k), but the centres of the discs do not. All discoids are bed-parallel; none exhibit evidence for folding, tearing, twisting, agglomeration in ripple troughs, or preservation indicative of a convex body (e.g., an exumbrella) that has been preserved upside-down. Sediment plugs (Fig. 4a–c, j, l, m) occur in approximately half of the specimens at the McKay Highway outcrop, and are typically ~2 cm in diameter; the largest plug is 8.75 cm in diameter. Of the 54 sediment plugs measured, 30 are of the same grain size as the adjacent sediment of the discoid; 18 of the plug-bearing specimens have finer sediment in the plug, and 2 have coarser sediment plugs.
15 Sectioning of 2 plugs and examination of edges of several broken slabs revealed no vertical shaſt below the plug surface (Fig. 5). For example, in Fig. 5a–c horizontal to inclined lami-nation occurs below the plug. In the central part of the discoid, a 1–3-mm-thick structureless mass of sediment occurs at the surface, corresponding to the area represented by the inner "ring" of the discoid (see arrows in Fig. 5). Flat-topped "plugless" discoids (e.g., Fig. 4g, h) also have horizontal to inclined lamination below their surface (Fig. 5e, f), and have no vertically oriented sediment or fluid shaſt below the surface.
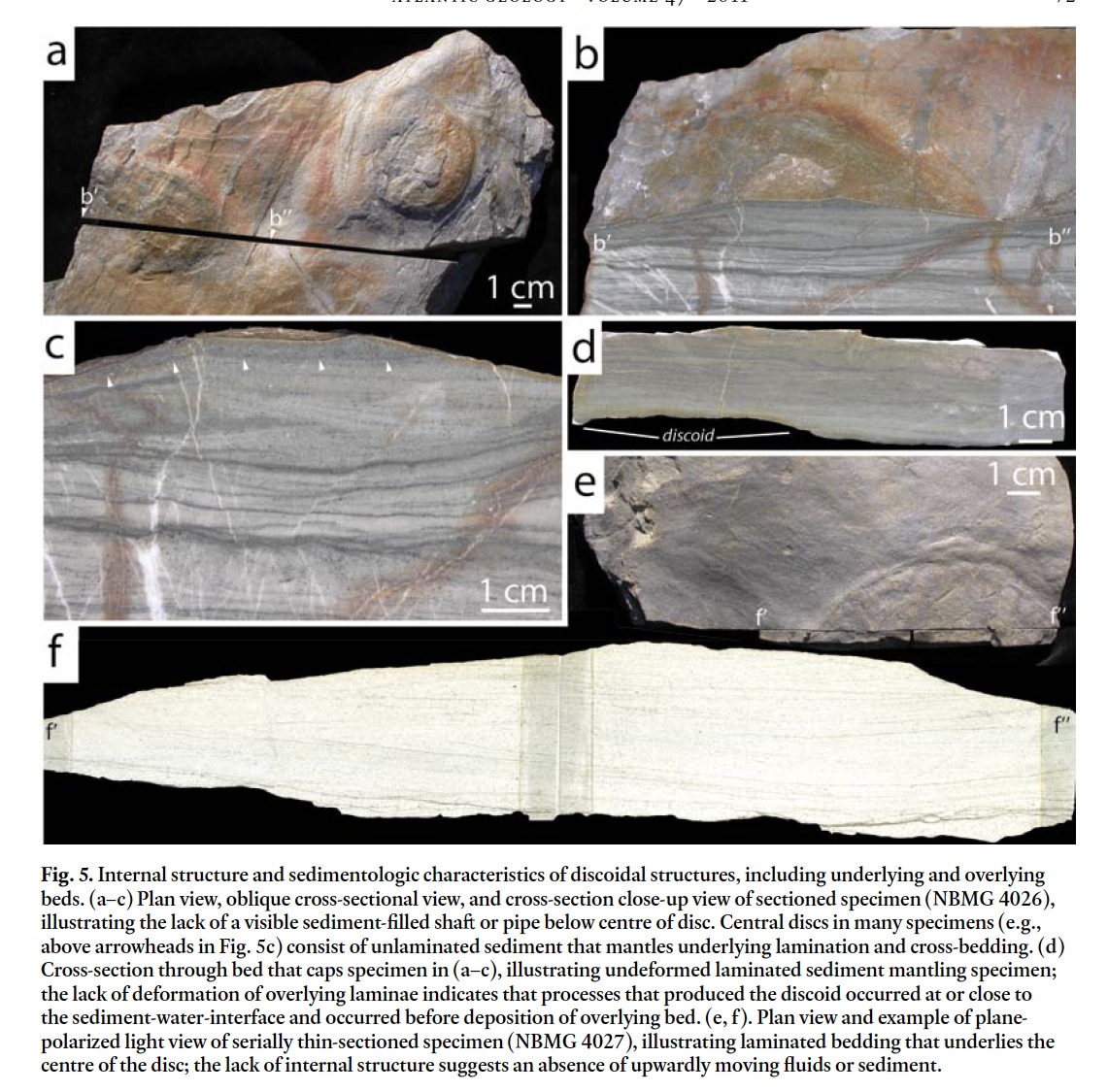 Display large image of Figure 5
Display large image of Figure 5
INTERPRETATIONS
16 Discoidal structures occur on at least three horizons (two of which are well exposed at the McKay Highway outcrop), and all share similar morphologic features, including features that are common to cnidarian medusae. Concentric rings, radial structures, bed-parallel orientation, and disc-like shapes are common in chondrophorine and related medusa (Hyman 1940; Fryer and Stanley 2004). Concentric rings, weakly disc-like shapes, radial structures (including radial cracks), a range of taphonomic grades (from well- to poorly preserved) and peripheral troughs are common features of fossil scyphozoan medusae (Schäfer 1941, 1972; Müller 1984; Bruton 1991; Young and Hagadorn, 2010).
17 Yet in all known deposits which bear abundant medusae, they oſten overlay, overlap, and deform one another — both when they are preserved bed-parallel, and when they are agglomerated in ripple or channel troughs (e.g., Haeckel 1866, 1869, 1874; Hagadorn et al. 2002; Hagadorn and Belt 2008; Young and Hagadorn 2010). Depending on the shape of the bell and the length of the oral arms, some medusae are nearly always preserved exumbrella down (Schäfer 1941; Kornicker and Conover 1960; Hertweck 1966; Bruton 1991). None of the King Square discoids exhibit these features, even among the few discoids that touch one another.
18 The radial structures on the discoids are not consistent in number, size, shape, or arrangement as one might expect among a similarly sized population of cnidarian medusae. Cross-cutting of outer rings by radial structures is atypical of stranded or fossilized scyphozoan medusa, because their radial structures are internal or dorsal features of their umbrellas. Moreover, the radial structures of the King Square specimens vary greatly in length within the same specimen — and although variance in such structures occurs in modern medusae, radial canals and comparable structures alternate in length in an alternating or a 2n pattern (e.g., Hyman 1940). The concentric rings of the King Square specimens are not usually circular or parallel to one another as in chondrophorine floats; rather, they exhibit weak semi-parallel arrangement and oſten have a sinuous to wavy pattern (e.g., Fig. 4a, c, j, l). Unlike extant medusae, which bear concentric structures, the King Square concentric rings are not consistent in number among specimens of similar size (Appendix 1) or among comparably preserved individuals.
19 In addition to identifying the presence of radial canals, Pick-erill (1982) hypothesized that some specimens preserved the outline of a stomach and a central mouth-like structure. In contrast, we observed no stomach or mouth, nor were able to observe reproducible structures that might be interpreted as gonads, tentacles, lappets or oral arms. No evidence of folding, tearing, shearing, twisting, or dragging is visible in the discoids or on bed surfaces across which they might have been dragged during deposition. Yet if these were unattached fossils, they would exhibit evidence for transport; even when stranded on interference rippled surfaces, cnidarian medusae comparable in size to the King Square Formation discoids should preferentially collect in the ripple troughs — but they do not.
20 Considered together, the morphology and sedimentological context of the King Square Formation discoids are inconsistent with their interpretation as cnidarian medusae. The lack of twisting and agglomeration , their bed-parallel orientation, and the internal bedding also rules out their origin as other mobile or transported organisms. It points to an origin in or on the sediment. Yet the lack of vertical shaſts or pipes of sediment in plug-bearing discoids is inconsistent with origin in the sediment by an animal’s burrowing activities or by an animal maintaining an open shaſt. The King Square structures are also unlike plug-shaped burrows such as Bergaueria (Pemberton et al. 1988) or stellate resting or escape traces (Häntzschel 1970). The lack of plugs in many specimens likewise precludes their origin as a vertically-oriented burrow or as an escape trace by an infaunal or epifaunal animal like an anemone. The lack of radiating three-dimensionally-preserved lobes on bed soles precludes interpretation of the King Square structures as Ediacara-type fossils such as Cyclomedusa or Eoporpita. The lack of consistent orientation of central plugs or evidence of tugging or current alignment typical of holdfasts of charniodiscids in the Rawnsley Quartzite or Mistaken Point Formations (South Australia and Newfoundland, respectively; Jenkins and Gehling 1978; Peterson et al. 2003; LaFlamme et al. 2004) precludes origin of these King Square structures as sheared-off disc-shaped holdfasts of Ediacaran fronds.
COMPARISON WITH OTHER SEDIMENTARY STRUCTURES
21 The most parsimonious interpretation for the King Square Formation discoids is that they are inorganic sedimentary structures produced by upward doming and subsequent cracking or bursting of a surface or near-surface bed. The largest sediment plugs in these structures are comparable to some sand volcanoes known from coarser turbidite sequences (Allen 1984). However, in light of the compositional and sedimentologic variation in central plugs and the lack of vertical shaſts below our sectioned plug-bearing specimens, it seems unlikely that these are persistent mud or sand volcanoes. Rather, they may be short-lived blisters and fluidization structures produced by dewatering or degassing of underlying sediment or organic matter. We found no positive evidence for the presence of a circular or round carcass buried below the surface, yet we cannot falsify the possibility that these structures originated through burial of such an organism. Some expelled sediment is occasionally visible, as demonstrated by the 1–4- mm-thick veneer of sediment that mantles the centre of discoid in Fig. 5c; and as suggested by the troughs surrounding several plug-bearing discoids, which could originate from collapse of adjacent or underlying sediment expelled through the plug of the discoid. However, many discoids lack accumulated sediment atop and around their margins.
22 Astropolithon are radial to concentric, blister-like sedimentary structures formed through blistering and upward movement of water, gas, and/or sediment through horizontally bedded sediments; they range in size from several millimetres to tens of centimetres in diameter, are always on bed tops in convex epirelief, have distinctive radial and concentric ornamentation, and oſten bear central sediment plugs (see reviews in Häntzschel 1975; Pickerill and Harris 1979). Originally described as a trace fossil (Dawson 1878), Astropolithon are now regarded as inorganic sedimentary structures (Pickerill and Harris 1979), but may form in association with microbially bound sediment (Seilacher et al. 2002). Astropolithon are most common in turbidite or prodeltaic settings where rapid alternating deposition of oversaturated mud and sand occurs. Like the King Square examples, Astropolithon are commonly associated with load structures, convolute bedding, or evidence of sand or mud dike intrusion (see review in Allen 1984). Non-radiate Astropolithon are known from modern sea-floor environments, and form a continuum of disc-shaped to radial structures that include pit-and-mounds, patterned cones, and allied structures (Ricci Lucchi 1970; Reineck and Singh 1973; Bryhni 1976; Allen 1984; Collinson et al. 2006). Most of these structures are produced by fluidization and upward movement of sediment due to thixotropic changes in beds deposited under rapid rates of sedimentation (Walter 1972; Pickerill and Harris 1979; Allen 1984).
23 The sediment infill in the King Square radial structures suggests a similar origin to Astropolithon, but also suggests that the cracks in the upward-doming bed must have been held open long enough for sediment to fall in. This radial infilling could occur when the dome was still active, by sediment expelled from the central shaſt, or may occur if the upward doming of sediment occurred intrastratally, perhaps a few millimetres below the sediment water interface, whereby overlying sediment could fall into the radial cracks and hold them "open" aſter the dome or blister had collapsed and was later compressed. Seilacher et al. (2002) invoked the presence of microbial biofilms or mats to facilitate doming and cracking processes in the formation of Astropolithon (see also Dornbos et al. 2007), but we found no positive evidence of such biofilms in thin-sections or on bed interfaces of King Square structures.
IMPLICATIONS
24 Although they are not medusae, the King Square sedimentary structures are nevertheless rare and intriguing. They provide insights into blistering, cracking and fluidization processes operating at the sea floor nearly 500 million years ago. Clearly, the variation in the internal structure of plug- and plugless Astropolithon and their flat-topped versus radially cracked morphologies suggest that we do not completely understand how they formed. We know such features form today in deep-sea and prodelta settings, but yet such structures are scarce in the rock record, suggesting that they are not readily preserved. Thus, a logical next step is to further constrain the conditions under which they form and the conditions that facilitate their preservation, by conducting laboratory degassing, fluidization, and burial experiments.
 Display large image of Table 1
Display large image of Table 1
We thank M. Mildenberger and R. Pickerill for help in the field and with references. S. Jensen and R. MacNaughton provided very helpful reviews. JWH is grateful for financial support from a George Frederic Matthew Fellowship from the New Brunswick Museum.