Oligocene benthic foraminifera from the Paleogene Wenonah Canyon, Scotian Shelf – normal versus canyon assemblages
Francis C. ThomasNatural Resources Canada, Geological Survey of Canada (Atlantic), Bedford Institute of Oceanography, P.O. Box 1006, Dartmouth, Nova Scotia B2Y 4A2 Canada
fthomas@nrcan.gc.ca
Date received: January 15, 2004 ¶ Date accepted: October 29, 2004
ABSTRACT
Petro-Canada Shell Wenonah J-75 well was drilled at the site of the mid Cenozoic "Wenonah Canyon" under the Scotian Shelf. Benthic foraminiferal assemblages recovered from sidewall core samples in the Oligocene section are unusually low in species diversity, though high in numbers of specimens. Such oligospecific assemblages may be the result of unusual hydrological factors in the canyon sediments at the time of deposition. In marked contrast, a nearby well, Canterra et al. Whycocomagh N-90, drilled outside the deepest canyon area, contains normal, more diverse, benthic slope assemblages. A seismic section acquired near the Wenonah site shows the informally designated Ypresian Acadia chalk to be truncated by canyon development. Planktic foraminifera of early Ypresian age do occur in Wenonah J-75, however, suggesting that the Acadia chalk may in fact be confined to the upper Ypresian.RÉSUMÉ
Le puits Wenonah J-75 de Petro-Canada/Shell a été foré à l'emplacement du « canyon Wenonah » du Cénozoïque moyen, sous la Plate-forme Scotian. Les assemblages de foraminifères benthiques récupérés des échantillons de carottes latérales dans la section de l'Oligocène, présentent une diversité d'espèces exceptionnellement faible, malgré le nombre élevé de spécimens. Ces assemblages oligospécifiques pourraient être dus à des facteurs hydrologiques inhabituels dans les sédiments du canyon au moment du dépôt. À l'opposé tout à fait, un puits proche, le puits Whycocomagh N-90, de Canterra et coll., foré à l'extérieur du secteur le plus profond du canyon, renferme des assemblages de pentes benthiques plus diversifiés et normaux. Une section sismique prélevée près de l'emplacement de Wenonah révèle que la craie officieusement désignée craie yprésienne d'Acadia est tronquée par le développement du canyon. Des foraminifères planctoniques de la période de l'Yprésien précoce sont présents dans le puits Wenonah J-75, ce qui laisse toutefois supposer que la craie d'Acadia pourrait en fait être confinée à l'Yprésien supérieur.[Traduit par la rédaction]INTRODUCTION
1 Over 200 exploration and delineation wells have been drilled on the Scotian Margin since 1966. Many of these have been the subject of micropaleontological investigation, including over 20 with data and interpretations concerning the Cenozoic interval (Ascoli 1976; Thomas 2002). The Cenozoic benthic foraminiferal assemblages in most of these wells, as described from cuttings samples plus occasional sidewall core samples, are typical for neritic or upper or lower bathyal conditions. They usually reflect a net regression from bathyal conditions in the early Paleogene to a shallower, neritic setting by the late Neogene. An extensive bibliography of published and unpublished data on the micropaleontology of these wells is given in Thomas (2002).
2 Cenozoic benthic assemblages on the Scotian Margin are generally rich and diverse, commonly with 20 or more foraminiferal species per sample (Ascoli 1976; Gradstein and Agterberg 1982; Thomas 2002 and unpublished data). However, at some levels in some wells, diagenesis, rapid sedimentation, unfavourable bottom conditions or other factors limit assemblages to low numbers of a few species: nevertheless, a moderate diversity of species is retained in all but the poorest samples. In some cases calcareous forms are nearly or entirely absent, replaced by agglutinated assemblages, which may be rich and diverse. These trends are evident from examination of cuttings samples, even though the nature of such samples precludes detailed statistical counts (see Materials and Methods). In only a few cases have discrete, unmixed benthic assemblages of Cenozoic age been recovered from east coast wells, providing rare opportunities for paleoecological study (Thomas 1991).
3 In one well however, Petro-Canada-Shell Wenonah J-75, examination of sidewall cores from the Oligocene interval has revealed unusual, oligospecific assemblages of benthic foraminifera. Significantly, seismic records indicate that this well was drilled in a site that was, during much of the mid Cenozoic, the location of a deep canyon (informally termed "Wenonah Canyon" by R.A. MacRae, personal communication 2001) that debouched onto the slope. Wenonah Canyon is approximately 11 km across at its widest point, and its maximum depth, as inferred from seismic data, is approximately 475 m (uncorrected for compaction). At its greatest development, in the mid Oligocene, its actual depth may have been some 500– 600 m (J. Shimeld, personal communication).
4 The purpose of this paper is to compare these unusual assemblages with coeval, "typical" benthic foraminifera from a nearby well, Canterra et al. Whycocomagh N-90, situated in a shallower setting, closer to a normal shelf environment (Thomas 2001b; Fig. 1).
Fig. 1 Location map showing well sites, seismic line 83-4411a, and areal extent of Wenonah Canyon, as inferred from regional seismic studies (A. MacRae, personal communication, 2002).
MATERIALS AND METHODS
5 All cuttings samples used in this study were prepared by the Canada-Nova Scotia Offshore Petroleum Board (CNSOPB), and picked by Atlantic Paleo Services of Dartmouth, Nova Scotia, in the late 1970s using methods described in Thomas (2002). The sidewall core samples (SWC's) were prepared and picked by or for the well owner, and processing details are not available. Selected specimens were imaged by the author using an ElectroScan E3 Environmental Scanning Electron Microscope, housed at the Geological Survey of Canada (Atlantic).
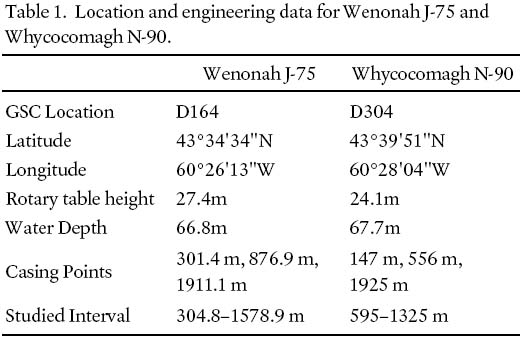
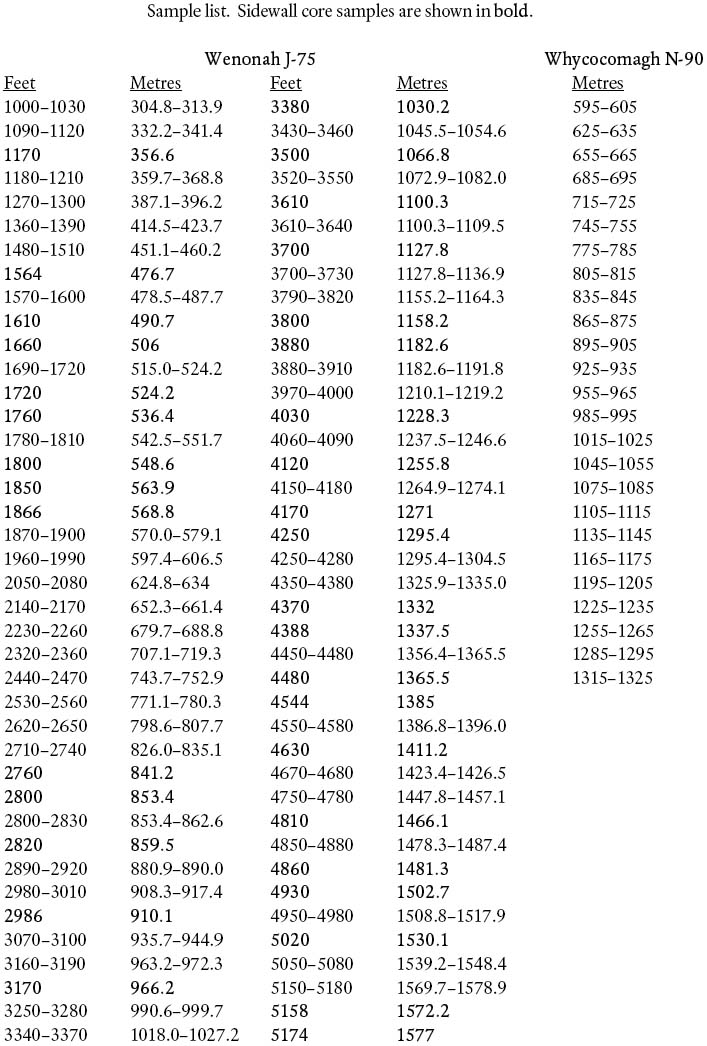
6 Note that, unlike Whycocomagh N-90, Wenonah J-75 was drilled prior to metrification, and depths on all samples and slides stored both at CNSOPB and at Geological Survey of Canada (Atlantic) were recorded in feet. All depths have been converted to metres for the purposes of this paper. Location and engineering data for both sites are provided in Table 1. Depths of all samples from both wells used in this study are given in Appendix 1.
Wenonah J-75
7 Petro-Canada-Shell Wenonah J-75 was spudded September 26, 1976, as an exploratory well on the Sable Subbasin, Scotian Shelf, about 30 km northwest of the present shelf break and 40 km southwest of Sable Island. The Cenozoic micropaleontology of this well was first studied by Gradstein (1978), but at that time only cuttings samples were available. The present author re-examined the cuttings (Thomas 2001a) along with the suite of 33 sidewall core samples that have become available since Gradstein's study.
8 Seismic records show that this well is situated near the axis of the informally named Wenonah Canyon, which developed on the ancient Scotian Slope as early as the Ypresian or, more likely, Rupelian (R.A. MacRae, personal communication 2001). The canyon was largely infilled by late Miocene (Tortonian) time, but for most of its existence it was subject to a distinct and much deeper depositional regime than would be expected given its distance and location inshore from the shelf break (Figs. 1 and 4).
9 The infilling of the canyon from the Rupelian on may simultaneously explain the thick Oligocene and Miocene sections in Wenonah J-75, the paucity of planktic foraminifera at most levels, the poverty of the benthic assemblages at some levels, and the unusual, oligospecific assemblages in others.
Table 1. Location and engineering data for Wenonah J-75 and Whycocomagh N-90.
Display large image of Table 1
10 A palynological (mainly dinoflagellate) biostratigraphic study of Wenonah J-75 (R.A. Fensome, 2003) is in general accord with the micropaleontological data for the Cenozoic section, particularly in the Paleogene, though some disparities exist (Fig. 2).
Fig. 2 Comparison of foraminiferal and dinoflagellate Cenozoic biostratigraphy for Wenonah J-75.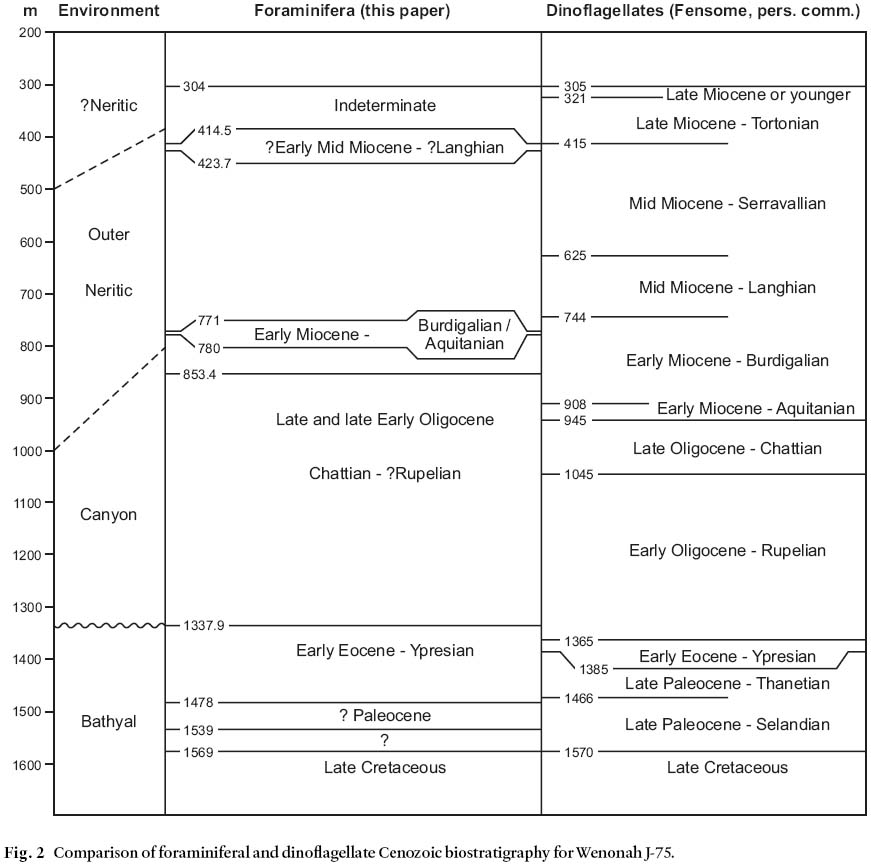
Display large image of Figure 2
11 The top of the Cretaceous section in Wenonah J-75 (Fig. 3) was determined by a sidewall core sample from 1365.5 m containing abundant Guembelitria cretacea, a Late Maastrichtian planktic foraminifera. Cuttings samples, in contrast, contain principally Lutetian and Ypresian benthics and planktics all the way down to 1569.7 m, at which point these caved forms are outnumbered by Cretaceous taxa. This is a clear demonstration of the frequent superiority of sidewall core samples over cuttings samples for detailed biostratigraphic analysis.
Fig. 3 Foraminiferal biostratigraphy of the Cenozoic section of Wenonah J-75.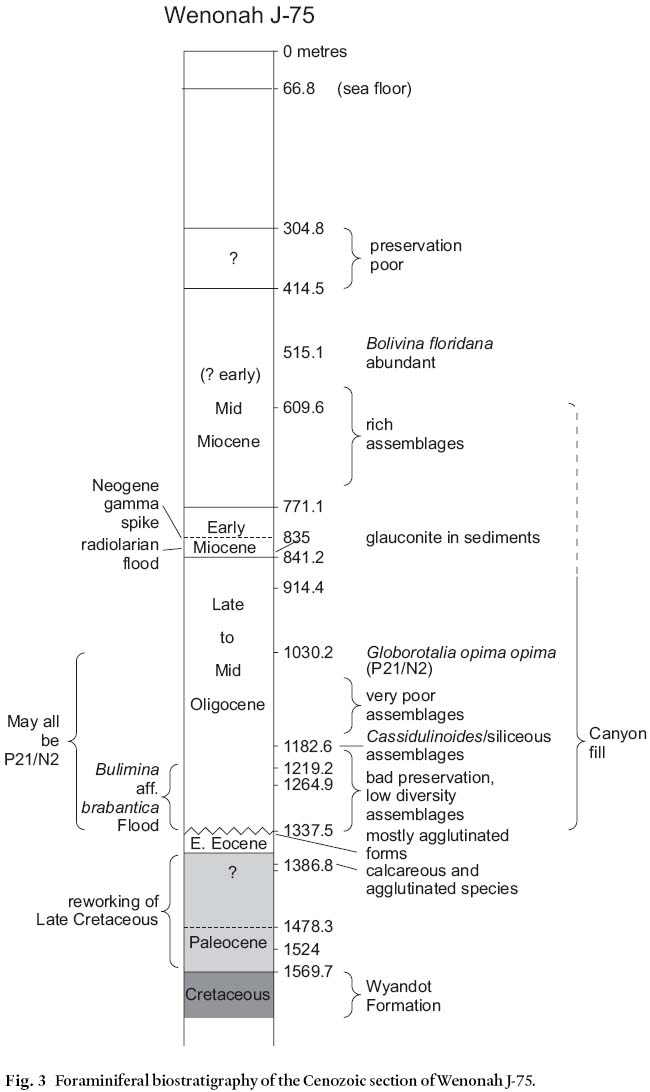
Display large image of Figure 3
12 Above the top Cretaceous, the interval 1365.5– 1337.5 m, represented by one cuttings sample and one sidewall core sample, contains a largely agglutinated Ypresian assemblage with rare planktic foraminifera and a few bathyal calcareous benthic taxa, usually poorly preserved. Many of these benthic species are found only in the early Ypresian of other Scotian Shelf sites (Thomas, unpublished data). Poor preservation of this carbonate material and the proliferation of agglutinated species suggests bottom water low in dissolved CaCO3. A few additional planktic species of this age, including Morozovella aequa, a diagnostic marker for the early Ypresian, are seen only as cavings in cuttings at 1396.0– 1386.8 m, well below the K/T boundary.
13 A laterally-extensive chalk unit of Ypresian age, informally named the Acadia chalk by industry operators in other parts of the Scotian Shelf (R.A. MacRae and J. Shimeld, personal communication 2003), is evidently eroded away by canyon development at the Wenonah site. Since early Ypresian micro-fossils are present in Wenonah even though the Acadia chalk is not, it follows that this unit may in fact be late Ypresian in age. Seismic line 83-4411a (Soquip project 8620-5014-006E, Fig. 4) crosses the canyon near Wenonah J-75, and clearly illustrates the truncation of the Acadia chalk.
Fig. 4 Seismic Line 83-4411a, showing erosional surface of Wenonah Canyon ( _ _ _ _ ), the Acadia chalk (----- ), and the top ofthe Wyandot Formation (. . . . .). Digital seismic data courtesy of the ex-Parex Group. (Early M. Mio = early Mid Miocene; E. Mio =Early Miocene; Oli = Oligocene; Ypr = Ypresian; Pal = Paleocene; LK = Late Cretaceous.)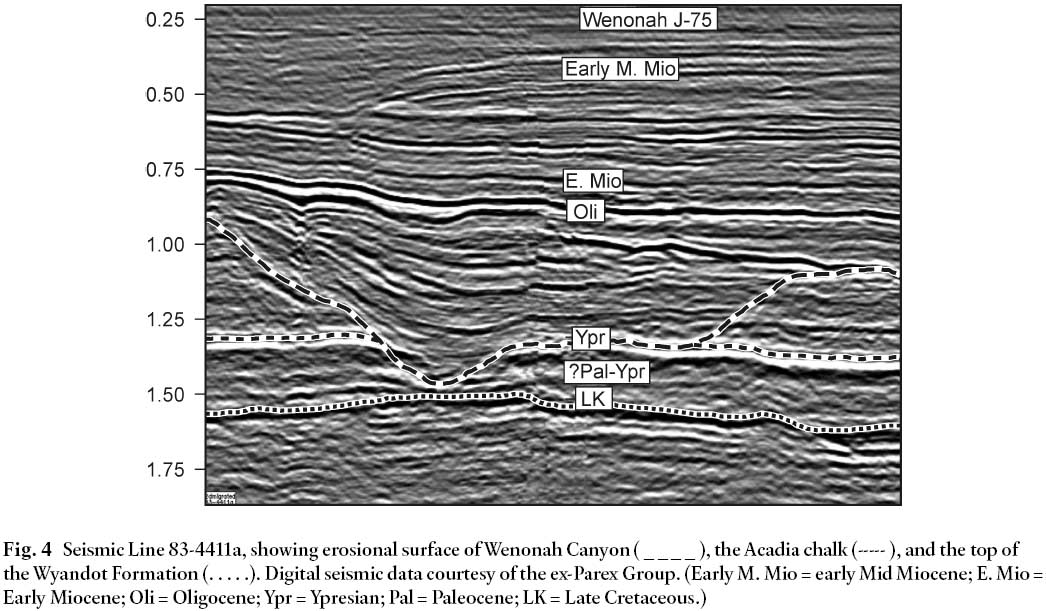
Display large image of Figure 4
14 The relatively thick (1335– 853.4 m) Oligocene section unconformably overlying the Ypresian exhibits a wide range of foraminiferal assemblages, differing in composition, diversity, and preservation. In the lower half, many samples, particularly SWC's, show either very impoverished assemblages, or are barren, whereas others contain large numbers of benthic foraminifera, but representing only two or three different species. Table 2 summarizes the microfossil content of the SWC samples.
Table 2. Microfossil content of sidewall core samples, Wenonah J-75.
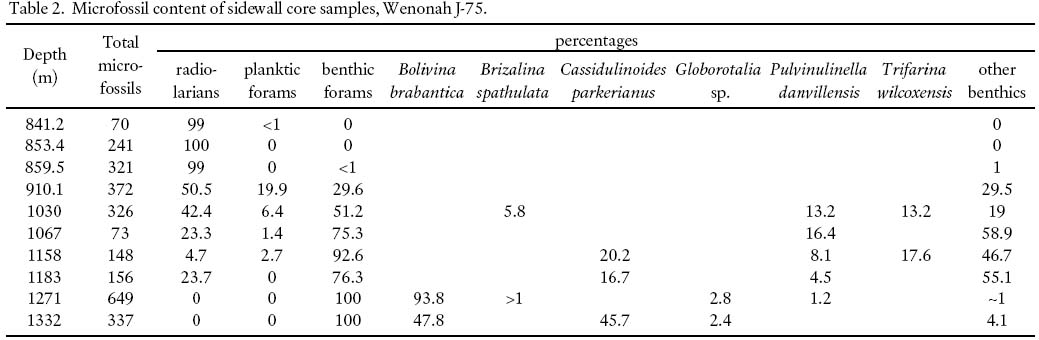
Display large image of Table 2
15 For most of the lower part of this interval, up to a cuttings sample at 1191.8– 1182.6 m, preservation of calcareous foraminifera is very poor. Most specimens occur only as silicified casts; they are often broken, flattened, or otherwise distorted. Above this level, from 1158.2 to 1066.8 m, samples are almost entirely barren. At 1030.2 m, a sidewall core sample shows a somewhat richer assemblage with a number of specimens of the planktic foraminifera Globorotalia opima opima, allowing for firm dating of this level as mid Oligocene (P21/N2). The next 150 m, up to the Miocene section, again contain generally impoverished assemblages.
16 The infilling of the canyon at the Wenonah site during the Oligocene may account for the episodically impoverished foraminiferal assemblages in two ways. Firstly, high rates of sedimentation tend to dilute benthic microfossil assemblages; and secondly, sudden influxes of sediment load in debris flows or turbidites can kill off and bury established benthic communities. It is possible that some of the near-monotypic assemblages seen in the Oligocene section may represent "pioneering" situations where fresh substrate after a depositional event provided an ideal habitat for only a few species until a more diverse fauna could become established.
17 Near the top of the Oligocene section, the sidewall core sample at 841.2 m contains a high concentration of glauconite. In this sample and in a few others just above and below it, abundant radiolarians comprise the entire microfossil suite. This association of glauconite and radiolarians is unusual – glauconite, if formed in situ, is classically thought to represent shallowing depositional environments, whereas large numbers of radiolarians are usually considered to represent deep, pelagic conditions. In many instances where radiolarians are well preserved, however, it is often taken to indicate that deposition was very rapid, protecting the relatively unstable opaline silica of their skeletons from dissolution; this is the case, for example, in the Early Eocene of the Jeanne d'Arc Basin, offshore Newfoundland (Thomas 1995). It could be inferred that similar rapid deposition, at least episodically, accounts for the preservation of radiolarians in Wenonah Canyon. The glauconite may have been a constituent of sediments slumped down the canyon from shallower source areas on the shelf. This mineral is commonly encountered in Oligocene samples from Scotian Shelf wells (Thomas, unpublished data). Sorting during transport may have concentrated the glauconite grains prior to their deposition at the well site.
18 The Aquitanian/Burdigalian section (835.1– 771.1 m), apparently conformably overlying the Oligocene, contains moderately rich, diverse and fairly well-preserved calcareous benthic foraminiferal assemblages but few planktics.
19 The thicker Langhian/Serravallian interval (771.1– 414.5 m) may actually contain some earlier Miocene sedimentation; age-diagnostic planktic foraminifera are rare, and caving is extensive in this section. Rich assemblages of benthics typify these sediments, often dominated by Bolivina aff. -1278?oridana, Bulimina cf. elongata, Florilus pizzarense and especially Lenticulina spp.
20 Above this section lies a short interval (414.5– 304.8 m) of sediments containing a low diversity of benthic foraminiferal taxa, most of which could be of Miocene or Pliocene age, with a few species possibly suggesting an age as young as Pleistocene.
Whycocomagh N-90
21 Canterra et al. Whycocomagh N-90 was drilled as an exploratory well in the Scotian Basin, and is located on the outer Scotian Shelf, some 50 km southwest of Sable Island and 10 km north of Wenonah J-75. A set of cuttings samples, based on 10 m sections spaced 20 m apart, were used to define the biostratigraphic subdivision of the Cenozoic interval of this well (Fig. 5).
Fig. 5 Foraminiferal biostratigraphy of the Cenozoic section of Whycocomagh N-90.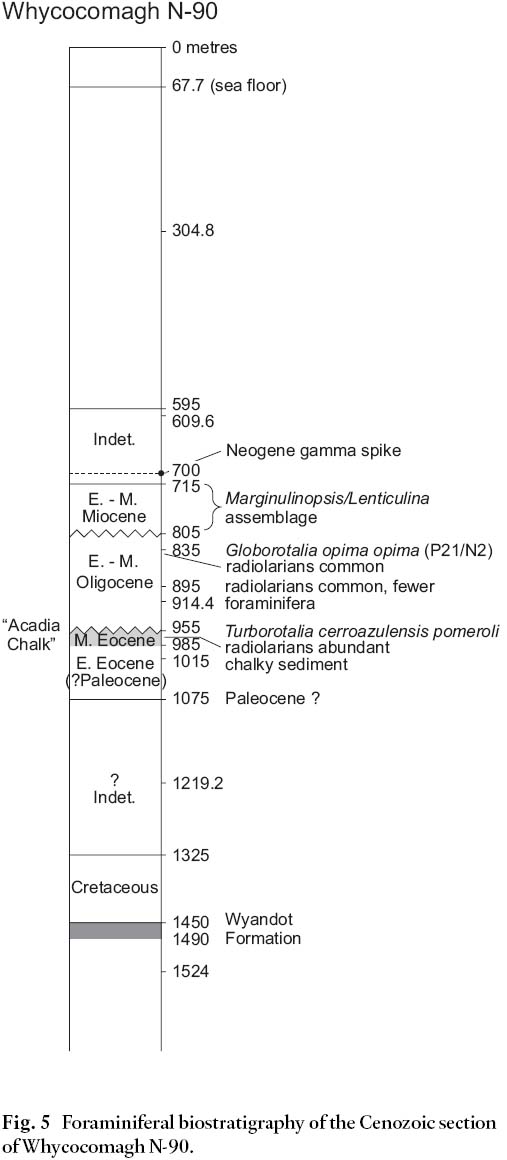
Display large image of Figure 5
22 As in the analysis of the Wenonah J-75 material, the generalized environmental interpretations are based largely on ratios of benthic to planktic foraminifera, and are usually corroborated by the types of benthic foraminifera or other taxa present.
23 The environment of deposition at the Whycocomagh site was bathyal from at least the Ypresian until the Oligocene, when shallowing conditions reduced the diversity and numbers of planktic foraminifera. The almost entirely benthic assemblages containing numbers of large, robust lenticulinids and marginulinids from the Miocene and at least part of the Oligocene are typical of shelf or neritic environments from Scotian Margin sediments of similar age. Regression continued into the Miocene, resulting in foraminiferal assemblages typical of lower neritic or shelf conditions.
24 The upper Rupelian interval (935– 805 m), which partially correlates with the Oligocene canyon interval of Wenonah J-75, contains a number of taxa common in coeval sediments in other parts of the Scotian Shelf (Thomas, unpublished data), with a few specimens of Globorotalia opima nana and Globorotalia opima opima providing reliable dates, if in place. The common radiolarians in this section are, as in the Wenonah site, somewhat unusual. In general the planktic:benthic ratio and the types of benthics suggest an upper slope environmental setting.
25 The preservation of calcareous microfossils in the Oligocene interval of Whycocomagh is moderately good, with most foraminiferal specimens stained buff, and many showing some degree of abrasion or breakage, suggesting a certain amount of transport and redeposition. The relatively rich planktic assemblages in this interval indicate that the Chattian and possibly some of the Lower Miocene is entirely missing.
DISCUSSION
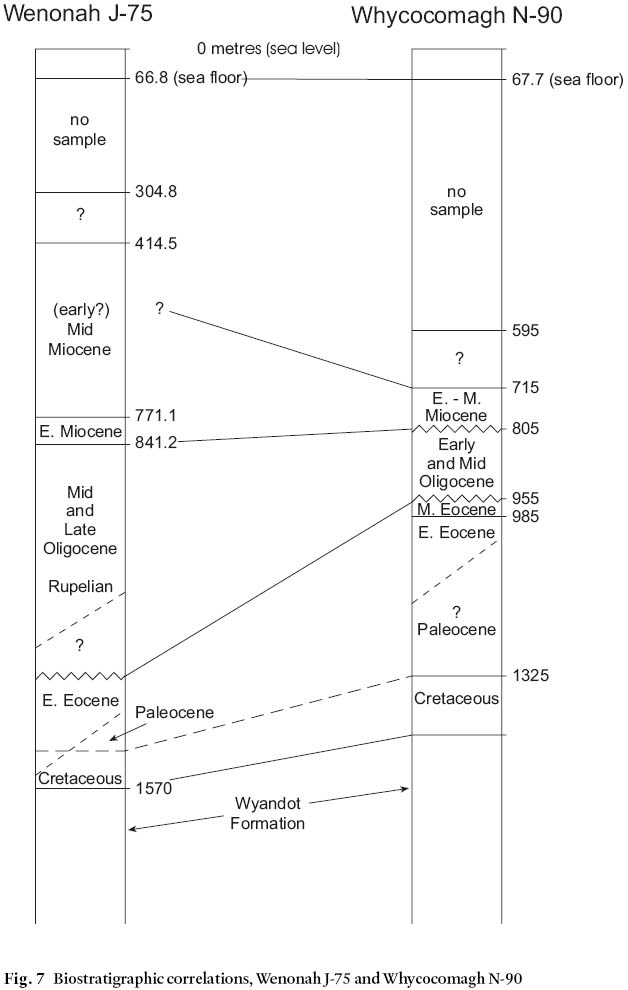
26 The oligospecific assemblages in the Wenonah sidewall cores are distinct from any others so far encountered in Cenozoic Canadian Atlantic offshore sites and it seems reasonable to ascribe this uniqueness to unusual sedimentological/chemical conditions in their original environments in the Wenonah Canyon. Some of the more common species in the sidewall core samples are illustrated in Fig. 6. The nearby Whycocomagh N-90 well, situated in a less incised area between canyon tributaries, contains no unusual assemblages. Figure 7 shows the Cenozoic biostratigraphic correlation between the two wells.
27 Studies reported in the literature clearly show that submarine canyons can contain benthic environments that are different from those in similar depths in normal slope environments. In a hydrologic study of currents in modern submarine canyons, Shepard et al. (1979) described alternating vertical currents of 25– 35 cm/sec, measured 3 m above the sediment surface. Because of their frequency and nature they were believed to be tide-related, and varied with depth along the axis of the canyons. They also reported short-lived turbidity currents flowing downhill at speeds of roughly 80 cm/sec.
28 Whether or not these special conditions in canyons always result in unusual benthic foraminiferal assemblages is a complex question, and the existing literature seems to contain a variety of answers. Weston (1985), in a study of modern canyon assemblages in piston cores in the eastern North Atlantic, found that canyons tend to support so-called "cosmopolitan" species (e.g. Brizalina dilatata andBrizalina subaenariensis); and furthermore, that normal depth stratifications of benthic species do not apply as well in canyons as in normal slope environments. Weston cited frequently changing water masses and variability of sediment supply as probable causes of these phenomena. In an examination of core samples from submarine canyons off the New Jersey coast, Jorissen et al. (1994) found that canyon samples tend to have relatively low benthic foraminiferal densities, and the infaunal suite of species is poorly developed, presumably due to sediment instability. These authors postulated that periodic small-scale mass wasting and bottom scour by currents may prevent benthic foraminiferal faunas from reaching a high-diversity equilibrium state. Furthermore, periodic shifts of the oxic-anoxic interface may tend to favour species with wider tolerance to low dissolved-oxygen levels. Vénec-Peyré (1990) found that, in canyons in the western Mediterranean, smaller foraminiferal taxa (both benthic and planktic) tend to be concentrated by hydrological factors near the canyon axes, and that diversity increased toward these axes.
29 In contrast, other studies of benthic foraminiferal assemblages in canyons, both modern and ancient, have found no particular anomalies. In an examination of material from Oligocene-Miocene canyon fill in west Africa, Petters (1982, 1984) reports only more or less "normal" benthic foraminiferal assemblages, in this case essentially the same mix of calcareous and benthic species found in contemporaneous deposits in nearby wells drilled on ancient slope environments. Similarly, Pícha (1979) reported no special benthic foraminiferal assemblages in Eocene-Oligocene canyon fill in central European sites, but did note the common presence of shallower-water taxa presumably carried down the canyon channel, and older forms supposedly eroding out of the canyon walls. It may be significant, however, that both of these studies relied almost exclusively on cuttings samples, which usually comprise material from a stratigraphic interval rather than a discrete horizon. In such samples, the characteristics of discrete samples from one instant in time can be totally masked by the addition of assemblages from adjacent levels.
30 There have been several studies of modern submarine canyons using samples from piston or box cores, or other relatively precise means, that have found no unusual benthic foraminiferal assemblages. Stanley et al. (1986) collected samples from the lower end of Wilmington Canyon, offshore Delaware, that contained "normal" benthic suites, although with a mix of shallower-water taxa ostensibly due to down-canyon transport. Petrological evidence suggests that deposition in the area was intermittent, and included a component of slumped sediment from eroding canyon walls. In a study of modern sediments from canyons off the Portuguese coast, Young et al. (1995) studied the accumulation rates of benthic foraminiferal tests. They found that these rates vary widely from canyon to canyon, and, though no especially high concentrations of these species were seen, the taxa Brizalina, Bulimina and the Cassidulininae had the highest overall accumulation rates. These taxa also figure prominently in the Wenonah samples.
31 In a later study of Wilmington and South Heyes canyons, Lundquist et al. (1997) found only more or less "normal" benthic assemblages, although several samples contained taxa indicative of shallower water as a result of downslope transport. They found no evidence, however, of high-energy downslope transport events in the last 200– 400 years, further suggestive of a stable benthic regime. An additional finding of this study was that the linearity of the canyon may affect the impact of bottom currents, with relatively straight stretches of canyon less affected than crooked ones.
32 Finally, Schmiedl et al. (2000) found that benthic foraminiferal faunas of canyons in the western Mediterranean do not generally contain opportunistic species, and are adapted to rather stable environments. In their study area there was a relatively large amount of phytodetritus and other organic matter as food sources, high standing benthic foraminiferal stock and diversity, a well developed deep infauna, and relatively high oxygen consumption rates. The open shelf areas at the bottom of the canyon actually showed lower standing stocks than the canyon samples. Possibly these assemblage characteristics, so different from the Wenonah material, may be attributable to the relative quiescence of the modern highstand, in contrast to the activity of the sea-level fall associated with the erosion of the Wenonah canyon during the mid Cenozoic.
33 While little or no relevant ecological data exists for most of the foraminiferal species in the Wenonah canyon assemblages, there are a few with close physical resemblances to modern forms. In such cases, while a direct lineal relationship with the Cenozoic form is conjectural, it is at least possible that the older form had similar environmental preferences to its modern counterpart.
34 For example, Trifarina wilcoxensis, which appears in relatively large numbers in two sidewall cores (1030.2 m and 1158.2 m) from the upper section of the canyon interval, may be an ancient analogue to modern Trifarina angulosa. The latter species has been listed as indicative of high-energy environments in areas off Antarctica (Violanti 1996), and one of two species (the other being Cibicides lobatulus) associated with strong bottom currents at depths of 200– 600 m on the Norwegian continental margin (Mackensen et al. 1985).
35 The globobuliminids, buliminids and bolivinids so common in some of the sidewall cores from the lower part of the canyon section may reflect reduced-oxygen conditions, as in the case described by Miller and Lohmann (1982) for the northeast United States continental slope. Furthermore, one bolivinid, Bolivina subaenariensis, has been identified with muddy, high-carbon sediments on the northern Florida continental slope by Sen Gupta et al. (1981). In this case, the sediments are from an area known for periodic local upwelling.
36 Finally, Cassidulinoides parkerianus, also common in the lower canyon interval, is one form which is still extant. While not widely distributed, it does appear to be more common at sites near continental margins, (Brady 1884; Matoba 1967; Ishman and Webb 1988) and so may have some affinity for areas of upwelling.
37 Possibly this succession of assemblages may reflect the evolution of the canyon, from a deeper-water environment subject to incursions of lower-oxygen bottom waters favouring the buliminids and bolivinids to somewhat shallower conditions with better oxygenation but more local dynamism in the benthic environment.
38 Significantly, the Oligocene interval of Whycocomagh N-90, some 10 km to the north, contains no unusual assemblages of benthic foraminifera (see Appendix 2); in fact none of the species occurring as important components of the sidewall core sample assemblages in Wenonah J-75 occur at all at Whycocomagh. Figure 7 illustrates the paucity of micropaleontological similarities between the two sites.
Fig. 6 (Below) (a, b, c, d, i) Bolivina aff. brabantica Kaasschieter. All specimens from sidewall core at 1271 m. (a) Length 0.314 mm, GSC No. 125291. (b) Length 0.356 mm, GSC No. 125292. (c) Length 0.318 mm, GSC No. 125293. (d) Length 0.458 mm, GSC No. 125294. (i) Apertural detail of same specimen. (e, f, g, h) Brizalina spathulata (Williamson). All somewhat damaged specimens from sidewall core at 1030.2 m. (e) Length 0.374 mm, GSC No. 125295. (f) Length 0.301 mm, GSC No. 125296. (g) Length 0.341 mm, GSC No. 125297. (h) Length 0.259 mm, GSCNo. 125298. (j, k, l) Trifarina wilcoxensis(Cushman and Ponton). All specimens from sidewall core at 1030.2 m. (j) Length 0.229 mm, GSC No. 125299. (k) Length 0.255 mm, GSC No. 125300. (l) Length 0.266 mm, GSC No. 125301. (m, n, o, p) Cassidulinoides parkerianus (Brady). All specimens from 1332 m. (m) Length 0.349 mm, GSC No. 125302. (n) Apertural detail of same specimen. (o) Length 0.363 mm, GSC No. 125303. (p) Length 0.554 mm, GSC No. 125304. (q, r, u, v) Pulvinulinella cf. danvillensis Howe and Wallace. All specimens from sidewall core at 1030.2 m. (q) Dorsal side, diameter 0.105 mm. Note circular dissolution pits. GSC No. 125305. (r) Dorsal side (note large dissolution pit in centre), diameter 0.121 mm, GSC No. 125306. (u) Ventral side, diameter 0.119 mm, GSC No. 125307. (v) Ventral side, diameter 0.110 mm (may have been slightly greater if undamaged), GSC No. 125308. (s, t) Unidentified spumellarian radiolarians from sidewall core at 1030.2 m. (s) Greatest diameter 0.249 mm, GSC No. 125309. (t) Greatest diameter 0.195 mm, GSC No. 125310.Fig. 7 Biostratigraphic correlations, Wenonah J-75 and Whycocomagh N-90.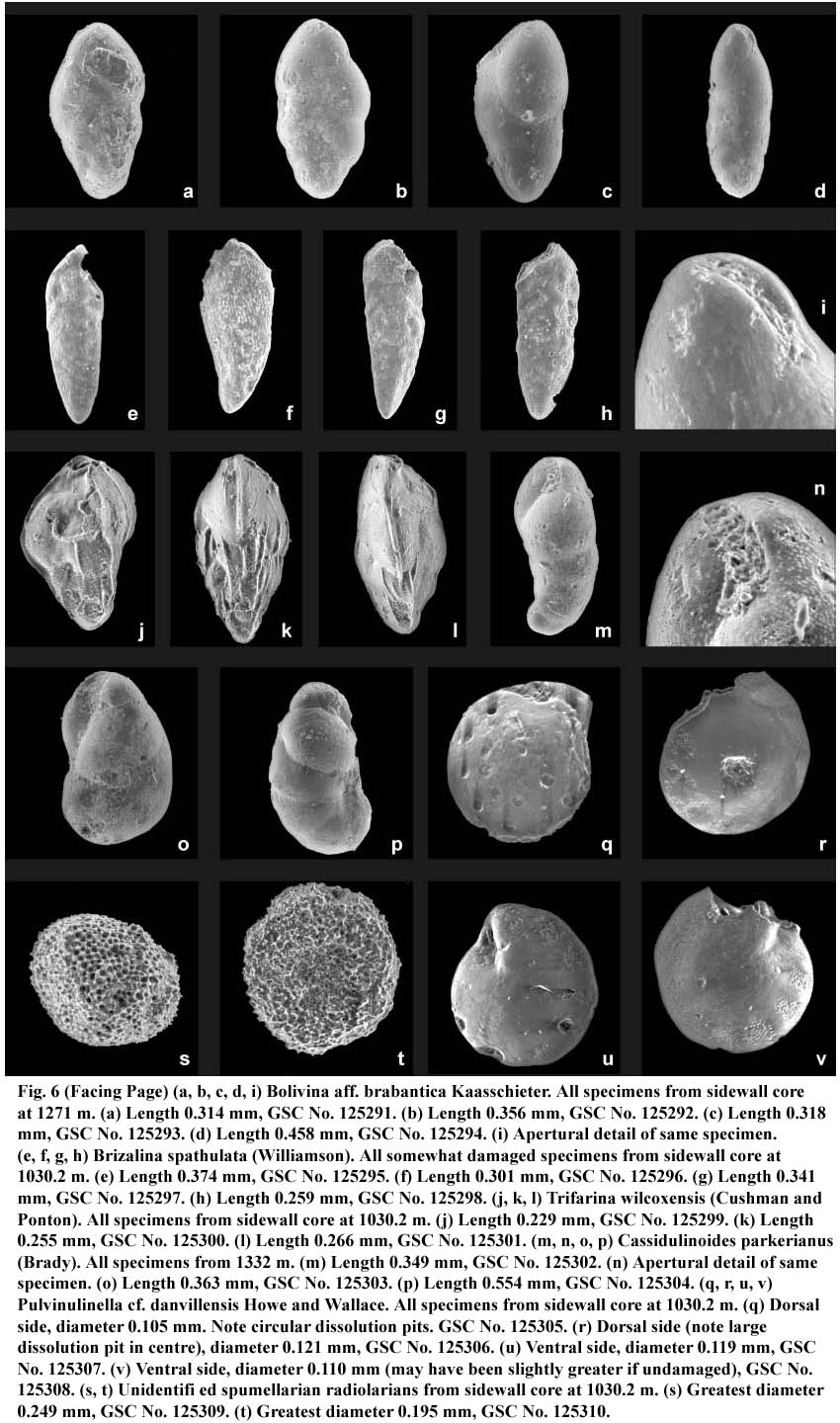
TAXONOMY OF SELECTED SPECIES
Bolivina aff. brabantica Kaasschieter, 1961 Fig. 6, a, b, c, d, i
aff. Bolivina brabantica Kaasschieter, 1961, p. 193– 194, pl. 8, fig. 22a, b.39 Remarks: Most specimens from Wenonah J-75 accord generally with Kaasschieter's original description of this Mid Eocene form in overall shape and relatively low numbers of chambers compared to many other bolivinids. The fine longitudinal striae he reports are barely visible on some specimens, though apparently absent on others. Some tend to have longer, less inflated chambers, presenting a slightly different, less lobate peripheral outline (Fig. 6c, d). This difference may relate to microspheric versus megalospheric forms. The generally poor preservation of the Wenonah material precludes positive assignment to Bolivina brabantica, a species that has not yet been noted from the Canadian Atlantic Margin.
40 In the Wenonah sidewall cores, this species is limited to two samples, at 1271 m and 1332 m (probable mid Oligocene).
Brizalina spathulata (Williamson 1858) Fig. 6, e-h
Textularia variabilis Williamson var. spathulata Williamson 1858, p. 76, pl. 6, fig. 164– 165.Bolivina ex. gr. spathulata (Williamson): Blanc-Vernet 1983, p. 504, pl. 1, fig. 2.
Bolivina spathulata (Williamson): Boltovskoy and Ocampo 1993, p. 147, pl. 1, fig. 7.
Brizalina spathulata (Williamson): Violanti 1994, p. 378. Brizalina spathulata (Williamson): Baggley 2000, pl. 1, fig. 17– 18.
41 Remarks: Although this species was originally described from Recent sands off Britain, its stratigraphic range has since been extended down into the Cenozoic. Boltovskoy and Ocampo (1993) found it in samples ranging in age from mid Oligocene to Pleistocene in the Arabian Sea, while Violanti's Swiss and Blanc-Vernet's Blake Outer Ridge specimens are of Pliocene age. Baggley (2000) reports it from the Miocene of Spain.
42 The Wenonah specimens greatly resemble Boltovskoy and Ocampo's figured specimen, particularly in the overall shape, and the lack of pores near sutures and down the central part of the test. Most specimens exhibit some breakage at the apertural end.
43 In Wenonah, Brizalina spathulata occurs in small numbers in two sidewall cores, at 1030.2 m (P21/N2 – Oligocene) and 1271 m (probable mid Oligocene).
Cassidulinoides parkerianus (Brady 1884) Fig. 6, m-p
Cassidulina parkeriana Brady 1881, p. 59; and Brady 1884, pl. LIV, fig. 11– 16.)Cassidulinoides parkerianus (Brady): Parr 1950, p. 344, pl. 12, fig. 25.
Cassidulinoides parkerianus (Brady): Matoba 1967, p. 253, pl. 28, fig. 23a, b, 24.
Cassidulinoides parkerianus (Brady): Ishman and Webb 1988, p. 534, pl. VI, fig. 1.
44 Remarks: Originally described from modern sediments, this characteristically "crosier-shaped" species (Brady 1881, ibid., p. 59) has since been widely reported: it has been found, for example, in the Pliocene of Japan (Matoba 1967) and Late Miocene to Late Pliocene intervals in the Antarctic (Ishman and Webb 1988). Wenonah specimens closely resemble all of these descriptions and figures, and are usually well preserved.
45 In the Wenonah material, Cassidulinoides parkerianus occurs in Oligocene sidewall cores at 1158.2 m, 1182.6 m and most abundantly at 1332 m. This species has not been previously noted from the Canadian Atlantic Margin, and these specimens apparently extend its worldwide known range into the Paleogene.
Pulvinulinella danvillensis Howe and Wallace 1932 Fig. 6, q, r, u. v
Pulvinulinella danvillensis Howe and Wallace 1932, p. 71, pl. 13, fig. 7a-c.46 Remarks: This small species has not previously been noted in Canadian Atlantic Margin sediments, possibly because routine microfossil picking has often excluded microfossils from this size range (less than 0.120 mm diameter). Note the circular dissolution pits shown in Fig. 6, q, r and u. It is fairly common in many of the Oligocene sidewall cores in Wenonah J-75.
Trifarina wilcoxensis (Cushman and Ponton 1932) Fig. 6, j, k, l
Pseudouvigerina wilcoxensis Cushman and Ponton 1932, p. 66, pl. 8, fig. 18a-b.Angulogerina wilcoxensis (Cushman and Ponton): Cushman and Garrett 1939, p. 84, pl. 14, figs. 24– 25. Angulogerina wilcoxensis Cushman and Garrett: Nogan 1964, p. 34, pl. 2, fig. 30.
Trifarina wilcoxensis (Cushman and Ponton): Bandy 1949, p. 145– 146, pl. 27, fig. 11a-b.
Trifarina wilcoxensis (Cushman and Garrett): Belanger and Berggren 1986, p. 336, pl. 3, fig. 5a-b.
47 Remarks: Originally described from Paleogene sediments in the southeastern United States, Trifarina wilcoxensis has been recorded in material as young as Early Miocene in the northeastern Atlantic (Belanger and Berggren 1986). Generally well preserved in the Wenonah samples, and common in two sidewall cores at 1030.2 m and 1158.2 m.
CONCLUSIONS
48 Seismic records indicate that the Wenonah J-75 well on the Scotian Shelf was the location, during the mid Cenozoic, of a tributary of a major submarine canyon incised into the shelf, debouching onto the lower slope.
49 Sidewall cores in the Oligocene section of Wenonah J-75 provide a unique opportunity to observe discrete benthic foraminiferal assemblages of Cenozoic age on the Scotian Margin. These samples contain unusual, oligospecific associations apparently related to the physiographic setting in the canyon. Unusual hydrographic conditions, especially at times during the Oligocene, may have led to the development of these associations, often dominated by relatively high percentages of just one, two or three species – taxa apparently capable of withstanding the unstable conditions in the canyon. As the canyon became more infilled and the depositional environment shallowed, these faunas gave way to more mixed benthic assemblages, often exhibiting breakage and other effects of a more dynamic environment. The coeval interval of nearby Whycocomagh N-90, a well in a non-canyon setting, contains no such assemblages.
APPENDIX 2
Biostratigraphic succession and faunal lists
Below are the faunal lists for each biostratigraphic unit encountered in the Cenozoic in Wenonah J-75, and in the Oligocene section of Whycocomagh N-90. Relative numbers of each taxon are denoted by: (ra) = rare; (co) = common; (ab) = abundant.
Wenonah J-75
Indeterminate 396.2– 304.8 m
Brizalina sp. (ra), Cibicides lobatulus (ra), Epistominella sp. (ra), Florilus pizzarense (ra) (?reworked), Globigerina sp. (ra), Islandiella cf. teretis (ra), Lenticulina cf. americana (co), Lenticulina calcarata (ra), Marginulina sp. (ra), Marginulina sp. (smooth) (co), Pyrgo sp. (ra), Quinqueloculina sp. (co)
Echinoid spines (co), fish bones (ra), Limacina sp. (pteropod) (ra), radiolarians (discoid), sponge spicules (co).
(?Early) Mid Miocene-?Langhian 752.9– 414.5 m
Arenobulimina sp. (ra), Asterigerina cf. gurichi (ra), Bolivina fl oridana (abundant at 515 m), Bolivina sp. (ra), Bulimina cf. elongata (ra, co at 661.4– 652.3 m), Bulimina aff. mexicana (ra), Bulimina sp. (ra), Cassidulina sp. (ra), Cassidulinoides sp. (ra), Catapsydrax dissimilis (ra), Cibicides lobatulus (ra), Cibicidoides cf. pachyderma (ra), Cibicidoides sp. 11 (ra), Epistomina umbonifera (ra), Florilus pizzarense (ab at 515.0 m, 661.4– 652.3 m), Globigerina bulloides (ra), Globigerina praebulloides (co), Globigerinita naparimaensis (ra), Globigerinoides trilobus (ra), Globoquadrina altispira (ra), Globoquadrina dehiscens (ra), Globorotalia continuosa (ra), Globorotalia cf. mayeri (ra), Globorotalia praemenardii (ra), Globorotalia scitula (ra), Globorotalia scitula-praescitula (co at 661.4– 652.3 m), Globorotalia sp. (ra), Globorotalia tumida tumida (ra, caved at 719.3– 707.1 m), Gyroidina girardana (ra), Gyroidina soldanii (co, ab at 661.4– 652.3 m), Lenticulina cf. americana (co, ab at 661.4– 652.3 m), Lenticulina iota (ra), Lenticulina vortex (ra), Marginulina aff. bachei (ra), Marginulina aff. decorata, (ra), Martinotiella communis (co), Melonis zaandami (ra), Nodosaria sp. (ra), Nodosaria sp. 1 (ra), Nodosaria sp. 4 (ra), Ophthalmidium sp. (ra), Planulina sp. (ra), ?Plectofrondicularia cf. barri (ra), Plectofrondicularia cf. vaughani (ra), Pullenia bulloides (ra), Quinqueloculina sp. (ra), Sphaeroidinellopsis seminula (752.9– 743.7 m), Spiroplectammina carinata (ra, co at 606.5– 597.4 m), Stilostomella sp., Textularia cf. agglutinans (co), Uvigerina canariensis (ra), Uvigerina peregrina (co, ab at 634– 624.8 m, 661.4– 652.3 m), Uvigerina sp. (small), Vulvulina sp. (ra).
Coscinodiscus sp. 1 (diatom) (ra), Coscinodiscus sp. 4 (ra), Coscinodiscus spp. (ab at 752.9– 743.7 m), echinoid spines (ra), fish otoliths (ra), mollusc fragments (ra), ostracods (ra), radiolarians (spherical, spindle-shaped) (co, ab at 752.9– 743.7 m), scaphopod sp. (ra, co at 688.8– 679.7 m), selachian denticles (ra), serpulid worms (ra).
Early Miocene-Aquitanian/Burdigalian 835.1– 771.1 m
Amphicoryna hirsuta (ra), Bathysiphon sp. (ra), Bulimina marginata (ra), Ceratobulimina contraria (ra), Cibicidoides sp. (co), Gaudryina sp. (ra), Globigerinoides trilobus (ra), Globocassidulina subglobosa (ra), Globoquadrina cf. dehiscens (ra), Guttulina problema (ra), Hoeglundina elegans (ra), Marginulina aff. decorata (ra), Martinotiella communis (ra), Melonis pompilioides (ra), Planulina wuellerstorffi (ra), Rectuvigerina striata (ra), Sigmoilinopsis schlumbergeri (ra), Trifarina sp. (ra), Uvigerina aff. proboscidea (ra).
Coscinodiscus spp. (ab at 780.3– 771.1 m), radiolarians (spherical, ab at 780.3– 771.1 m, 835.1– 826.0 m).
Late and mid Oligocene-?Rupelian/Chattian 1335.0– 853.4 m
Allomorphina aff. pauciloculata (ra), Anomalina sp. (ra), Biloculina sp. (ra), Bolivina aff. brabantica (co, ab at 1274.1– 1264.9 m, a sidewall core at 966.2 m, 1030.2 m), Brizalina spathulata (ab in sidewall cores at 966.2 m, 1030.2 m), Bulimina alazenensis (ra), Bulimina ovata (ra), Bulimina sp. (ra), Cassidulinoides parkerianus (co in sidewall core at 1158.2 m), Cassigerinella chipolensis (ra), Catapsydrax dissimilis (co), Cibicides sp. (co), Cibicidoides crebbsi (ra), Coarse agglutinated spp. (ra), Cyclammina sp. (ra), Epistomina umbonifera (ra), Fissurina marginata (ra), Globigerina praebulloides (ra), Globobulimina sp. (ra), Globoquadrina venezuelana (ra, co at 1335.0– 1325.9 m), Globorotalia opima-continuosa transition (co in sidewall core at 1030.2 m), Globorotalia opima nana (ra), Globorotalia opima opima (co in sidewall core at 1030.2 m, 1335.0– 1325.9 m), Melonis pompilioides (ra), Nodosaria sp. 11, Pullenia bulloides (co), Pullenia quinqueloba (ra), Pulvinulinella danvillensis (ab in sidewall core at 1030.2 m), Sphaeroidina bulloides (ra), Trifarina wilcoxensis (co), Turrilina alsatica (ra).
Fish otoliths (ra), fish teeth (ra), mollusc fragments (ra), radiolarians (co, ab in sidewall core at 1127.8 m).
Early Eocene-Ypresian 1365.5– 1337.5 m
Acarinina soldadoensis (ra), Adercotryma sp. (ra), Ammobaculites sp. (ra, caved to 1447.8 m), Ammodiscus planus (ra), Anomalinoides acuta (ra, caved to 1386.8 m), Bulimina bradburyi (ra, caved to 1386.8 m), Bulimina midwayensis (ra, caved to 1386.8 m), Bulimina trinitatensis (ra, caved to 1386.8 m), Cyclammina cancellata (ra), Epistomina cf. eocaena (ra), Gaudryina cf. laevigata (ra), Gavelinella beccariiformis (ra), Gavelinella danica (ra), Gavelinellopsis micra (ra), Globigerina linaperta (ra), Globorotalia intermedia-wilcoxensis, Gyroidina octocamerata (ra, caved to 1386.8 m), Glomospira corona (ra), Haplophragmoides kirki (ra), Karreriella cf. conversa (ra), Lenticulina whitei (ra), Morozovella aequa (ra, caved to 1386.8 m), Neoeponides sp. (ra, caved to 1386.8 m), Nuttalinella florealis (ra, caved to 1386.8 m), Planorotalites sp. (ra, caved to 1386.8 m), Pseudohastigerina micra-wilcoxensis (ra, caved to 1386.8 m), Recurvoides sp. (ra), Reticulophragmium amplectens (ra), Spirillina sp. (ra), Spiroplectammina dentata (ra, caved to 1478.3 m), Spiroplectammina navarroana (ra, caved to 1386.8 m), Spiroplectammina spectabilis (ra, caved to 1386.8 m), Subbotina patagonica (ra, caved to 1386.8 m), Tritaxia cf. aspera (ra), Trochamminoides sp. (ra, caved to 1386.8 m), Truncorotalites rohri (ra), Turrilina brevispira (ra, caved to 1478.3 m).
Fish teeth (ra), ostracods (ra), radiolarians (co, spherical, ovoid).
Late Maastrichtian 1365.5 m+
Guembelitria cretacea (ab in sidewall core at 1365.5m).
Whycocomagh N-90
Upper Lower to Mid Oligocene-Rupelian/?Chattian 805– 935 m
Catapsydrax dissimilis (ra), Cibicidoides sp. (ra), Eggerella sp. (ra), Eponides umbonatus (ra), Globorotalia aff. ampliapertura (ra), Globorotalia opima nana (ra), Globorotalia opima opima (ra), Gyroidina soldani (ra), Karreriella bradyi (ra), Lenticulina sp. (ra), Melonis pompilioides (co), Nodosaria sp. 4 (ra), Nodosaria sp. 14 (co), Nodosaria sp. 19 (co), Plectofrondicularia sp. (ra), Pullenia bulloides (ra), Pullenia quinqueloba (ra), Sphaeroidina bulloides (ra), Spiroplectammina sp. (ra), Uvigerina sp. (ra).
Fish tooth (ra), radiolarians (spherical, ovoid) (co).
ACKNOWLEDGMENTS
My thanks to: John Shimeld (GSC Atlantic), for valuable insights into the nature and timing of the Acadia chalk and providing the seismic section; Bill MacMillan (GSC Atlantic) for producing Fig. 6; Ken Hale (GSC Atlantic), for drafting the figures; Nelly Koziel (GSC Atlantic) for word processing and arranging the finished document; Andrew MacRae (Saint Mary's University) for his advice on the nature and timing of the Wenonah Canyon and critical review of the whole document, and Graham Williams (GSC Atlantic) for his continued support and encouragement. John Shimeld (GSC Atlantic) and Dave McNeil (GSC Calgary) reviewed the document and offered many helpful revisions, as did Rob Fensome (GSC Atlantic) in his role as editor for the journal. This is Geological Survey of Canada Contribution no. 2003219.REFERENCES
Ascoli, P. 1976. Foraminiferal and ostracod biostratigraphy of the Mesozoic-Cenozoic, Scotian Shelf, Atlantic Canada. In First International Symposium on Benthonic Foraminifera of Continental Margins, Part B: Paleoecology and Biostratigraphy. Edited by C.T. Schafer and B.R. Pelletier. Maritime Sediments, Special Publication 1, Halifax, Nova Scotia, pp. 653– 771.
Baggley, K.A. 2000. The Late Tortonian-Early Messinian foraminiferal record of the Abad Member (Turre Formation), Sorbas Basin, Almería, South-east Spain. Palaeontology, 43, pp. 1069– 1112.
Bandy, O.L. 1949. Eocene and Oligocene foraminifera from Little Stave Creek, Clarke County, Alabama. Bulletins of American Paleontology, 32, 210 p.
Belanger, P.E., & Berggren, W.A. 1986. Neogene benthic foraminifera of the Hatton-Rockall Basin. Micropaleontology, 32, pp. 324– 356.
Blanc-Vernet, L. 1983. Benthic foraminifers of Site 533, Leg 76 of the Deep Sea Drilling Project – faunal variations during the Pliocene and Pleistocene on the Blake Outer Ridge (Western North Atlantic). In Initial Reports of the Deep Sea Drilling Project LXXVI. Edited by R.E. Sheridan, F.M. Gradstein and others. United States Government Printing Office, Washington D.C., pp. 497– 509.
Boltovskoy, E., & Ocampo, J.V. 1993. Benthic foraminifers from DSDP Site 219 (Eocene-Pleistocene, Arabian Sea). Re-vista Española de Micropaleontologia, 25, pp. 127– 156. Brady, H.B. 1881. Notes on some of the reticularian Rhizopoda of the "Challenger" Expedition; Part III. Quarterly Journal of the Microscopical Society, London, New Series, 21, pp. 31– 71.
Brady, H.B. 1884. Report on the foraminifera dredged by H.M.S. Challenger during the years 1873– 1876, 9, 814 p. Cushman, J.A., & Garrett, J.B. 1939. Eocene foraminifera of Wilcox age from Woods Bluff, Alabama. Contributions of the Cushman Laboratory for Foraminiferal Research, 15, pp. 77– 88.
Cushman, J.A., & Ponton, G.M. 1932. An Eocene foraminiferal fauna of Wilcox age from Alabama. Contributions of the Cushman Laboratory for Foraminiferal Research, 8, pp. 51– 72.
Fensome, R.A. 2003. Palynological analysis of Petro-Canada-Shell Wenonah J-75 well, Scotian Shelf. Report No. MRGPAL. 5-2003RAF, 8 p. (Unpublished report.) Gradstein, F.M. 1978. Foraminiferal stratigraphy of Petro-Canada Shell Wenonah J-75. Report No. EPGS-PAL. 11-78FMG, 2 p. (Unpublished report.)
Gradstein, F.M., & Agterberg, F.P. 1982. Models of Cenozoic foraminiferal stratigraphy – northwestern Atlantic Margin. In Quantitative Stratigraphic Correlation. Edited by J.M. Cubitt and R.A. Reyment. John Wiley and Sons, Ltd., Chichester, U.K., pp. 119– 174.
Howe, H.V., & Wallace, W.E. 1932. Foraminifera of the Jackson Eocene at Danville Landing on the Ouachita, Catahoula Parish, Louisiana. Louisiana Department of Conservation, Geological Bulletin, 2, pp. 1– 118.
Ishman, S.E., & Webb, P.-N. 1988. Late Neogene benthic foraminifera from the Victoria Land Basin margin, Antarctica: Application to glacio-eustatic and tectonic events. In Benthos '86, Proceedings of the 3rd International Symposium on Benthic Foraminifera. Revue de Paléobiologie, Volume Spécial, 2, pp. 523– 551.
Jorissen, F.J., Buzas, M.A., Culver, S.J., & Kuehl, S.A. 1994. Vertical distribution of living benthic foraminifera in submarine canyons off New Jersey. Journal of Foraminiferal Research, 24, pp. 28– 36.
Kaasschieter, J.P.H. 1961. Foraminifera of the Eocene of Belgium. Institut Royal des Sciences Naturelles de Belgique, Mémoire 147, 271 p.
Lundquist, J.J., Culver, S.J., & Stanley, D.J. 1997. Foraminiferal and lithologic indicators of depositional processes in Wilmington and South Heyes submarine canyons, U.S. Atlantic continental slope. Journal of Foraminiferal Research, 27, pp. 209– 231.
Mackensen, A., Sejrup, H.P., & Jansen, E. 1985. The distribution of living benthic foraminifera on the continental slope and rise off southwest Norway. Marine Micropaleontology, 9, pp. 275– 306.
Matoba, Y. 1967. Younger Cenozoic foraminiferal assemblages from the Choshi District, Chiba Prefecture. Science Reports of the Tohoku University, Sendai, Second Series (Geology) 38, pp. 221– 263.
Miller, K.G., & Lohmann, G.P. 1982. Environmental distribution of Recent benthic foraminifera on the northeast United States continental slope. Geological Society of America, Bulletin 93, pp. 200– 206.
Nogan, D.S. 1964. Foraminifera, stratigraphy, and paleo-ecology of the Aquia Formation of Maryland and Virginia. Cushman Foundation for Foraminiferal Research, Special Publication 7, 50 p.
Parr, W.J. 1950. Foraminifera. British and New Zealand Antarctic Research Expedition Reports, ser. B, 5, pp. 232– 392.
Petters, S.W. 1982. Central West African Cretaceous-Tertiary benthic foraminifera and stratigraphy. Palaeontographica, Abteilung A, 179, pp. 1– 104.
Petters, S.W. 1984. An ancient submarine canyon in the Oligocene-Miocene of the western Niger Delta. Sedimentology, 31, pp. 805– 810.
Pícha, F. 1979. Ancient submarine canyons of Tethyan continental margins, Czechoslovakia. American Association of Petroleum Geologists, Bulletin 63, pp. 67– 86. Schmiedl, G., de Bovée, F., Buscail, R., Charrière, B., Hemleben, C., Medernach, L., & Picon, P. 2000. Trophic control of benthic foraminiferal abundance and microhabitat in the bathyal Gulf of Lyons, western Mediterranean Sea. Marine Micropaleontology, 40, pp. 167– 188.
Sen Gupta, B., Lee, R.F., & May, M.S. 1981. Upwelling and an unusual assemblage of benthic foraminifera on the northern Florida Continental Slope. Journal of Paleontology, 55, pp. 853– 857.
Shepard, F.P., Marshall, N.F., McLoughlin, P.A., & Sullivan, G.G. 1979. Currents in Submarine Canyons and other Seavalleys. American Association of Petroleum Geologists, Studies in Geology 8, Tulsa, Oklahoma, 171 p.
Stanley, D.J., Culver, S.J., & Stubblefield, W.L. 1986. Petrologic and foraminiferal evidence for active downslope transport in Wilmington Canyon. Marine Geology, 69, pp. 207– 218.
Thomas, F.C. 1991. Paleoecology of Late Eocene-Oligocene foraminiferal assemblages in a two-well transect across the northeast Newfoundland shelf. Journal of Micropalaeontology, 10, pp. 57– 67.
Thomas, F.C. 1995. A Paleogene radiolarian event of the South Mara unit, Banquereau Formation, Jeanne d'Arc Basin, offshore Newfoundland, and its applications. In Current Research 1995-E Geological Survey of Canada, pp. 211– 220. Thomas, F.C. 2001a. Petro-Canada-Shell Wenonah J-75 revisited – Cenozoic micropaleontology and paleoenvironments. Report No. MRG-PAL. 10-2001FCT, 9 p. (Unpublished report.)
Thomas, F.C. 2001b. Report on the Cenozoic biostratigraphy (Foraminifera) and depositional environments of the Canterra et al. Whycocomagh N-90 well (Scotian Shelf), from 595 to 1325 m (Cretaceous). Report No. MRG-PAL. 8-2001FCT, 6 p. (Unpublished report.)
Thomas, F.C. 2002. Geological Survey of Canada's Atlantic Margin Subsurface Micropaleontology Studies – History, Facts and Figures. Geological Survey of Canada Open File 4163, Ottawa, 55 p.
Vénec-Peyré, M.-T. 1990. Contribution of foraminifera to the study of Recent sedimentation in the Gulf of Lyons (western Mediterranean Sea). Continental Shelf Research, 10, pp. 869– 883.
Violanti, D. 1991. I Foraminiferi pliocenici di Castel di Sotto (Novazzano, Cantone Ticino): considerazioni biostratigrafiche e paleoambientale. Bollettino Societá Ticinense Scienze Naturali, 79, pp. 49– 70.
Violanti, D. 1994. Pliocene mud-dwelling benthic foraminiferal assemblages dominated by Globobulimina affi nis (Castel di Sotto, Canton Ticino, Switzerland). In Studies on Ecology and Paleoecology of Benthic Communities. Edited by R. Matteucci and others. Bollettino della Societá Paleontologia Italiana, Special Volume 2, pp. 365– 379. Violanti, D. 1996. Taxonomy and distribution of recent benthic foraminifers from Terra Nova Bay (Ross Sea, Antarctica), Oceanographic Campaign 1987/1988. Palaeontographica Italica, 83, pp. 25– 71.
Weston, J.F. 1985. Comparison between Recent benthic foraminiferal faunas of the Porcupine Seabight and Western Approaches Continental Slope. Journal of Micropalaeontology, 4, pp. 165– 183.
Williamson, W.C. 1858. On the Recent foraminifera of Great Britain. Royal Society of London, 107 p.
Young, M., Murray, J.W., & Croudace, I.W. 1995. Radionuclide dates and foraminiferal accumulation rates: examples from submarine canyons. Marine Micropaleontology, 26, pp. 57– 63.
Editorial responsibility: Robert A. Fensome