Article
Carboniferous palynoevents in the circum-Arctic region
doi:10.4138/atlgeol.2021.003
Abstract
The Carboniferous of the present-day Arctic yields an abundant and diverse association of terrestrial palynomorphs, particularly from Mississippian successions. Relatively few sections have been studied. However, those published so far demonstrate considerable similarities in the palynofloras between Arctic regions. Based on the published record, we present a compilation of 31 last occurrences (LOs), first occurrences (FOs), and some abundance events that have correlation potential around the Arctic. The chronostratigraphic resolution is relatively low, and the lack of independent age control hampers exact age assignments. But for most of these successions, palynology is the only dating tool available.
Résumé
Le Carbonifère de la région arctique d’aujourd’hui fait état d’une association abondante et diversifiée de palynomorphes terrestres, en particulier de successions du Mississippien. Relativement peu de sections ont été étudiées. Celles publiées jusqu’ici montrent toutefois des similarités considérables des palynoflores entre les régions arctiques. En nous basant sur les documents publiés, nous présentons une compilation des 31 dernières occurrences (DO), des premières occurrences (PO) et de certains phénomènes d’abondance qui pourraient être corrélés dans la région de l’Arctique. La résolution chronostratigraphique est relativement faible et l’absence de critères de datation gêne l’attribution d’âges exacts. La palynologie constitue toutefois le seul outil de datation disponible dans le cas de la majorité de ces successions.
[Traduit par la redaction]
INTRODUCTION
1 This article is a contribution to the Circum-Arctic Palynological Events (CAPE) project, providing a scheme of selected events for the Carboniferous in the present-day Arctic. The Carboniferous Period extended from 358.9 to 298.9 Ma according to Gradstein et al. (2020). The Carboniferous is divided into two subperiods, Mississippian and Pennsylvanian, the latter having a base at 323.2 Ma. The Mississippian is divided into Tournaisian, Visean and Serpukhovian stages, and the Pennsylvanian is divided into the Bashkirian, Moscovian, Kasimovian and Gzhelian stages. Historically, various chronostratigraphic schemes have been applied to the Carboniferous, making correlation and comparison between the various palynological zonations and charts difficult. However, as a “rule of thumb”, the previous use of Lower and Upper Carboniferous may be replaced by Mississippian and Pennsylvanian respectively, although the latter might include the latest Mississippian in some cases.
2 As palynomorphs are the most common fossil group recorded in Carboniferous strata, palynology is a critical tool for dating and refining the stratigraphic correlations of rocks of that period. This is particularly true for the Mississippian, a time during which non-marine clastics were the dominant deposits, a result of rifting in a series of local half-grabens in most Arctic areas. Carboniferous palynomorphs are therefore dominantly miospores deposited in fluvial and lacustrine sediments, although freshwater algae are sometimes abundant, and rare scolecodonts and marine acritarchs occur in the Visean. Palynological data is sparser for the Pennsylvanian successions in the Arctic, at a time when carbonate deposition dominated due to the shift from humid to arid climatic conditions and ongoing sea-level rise.
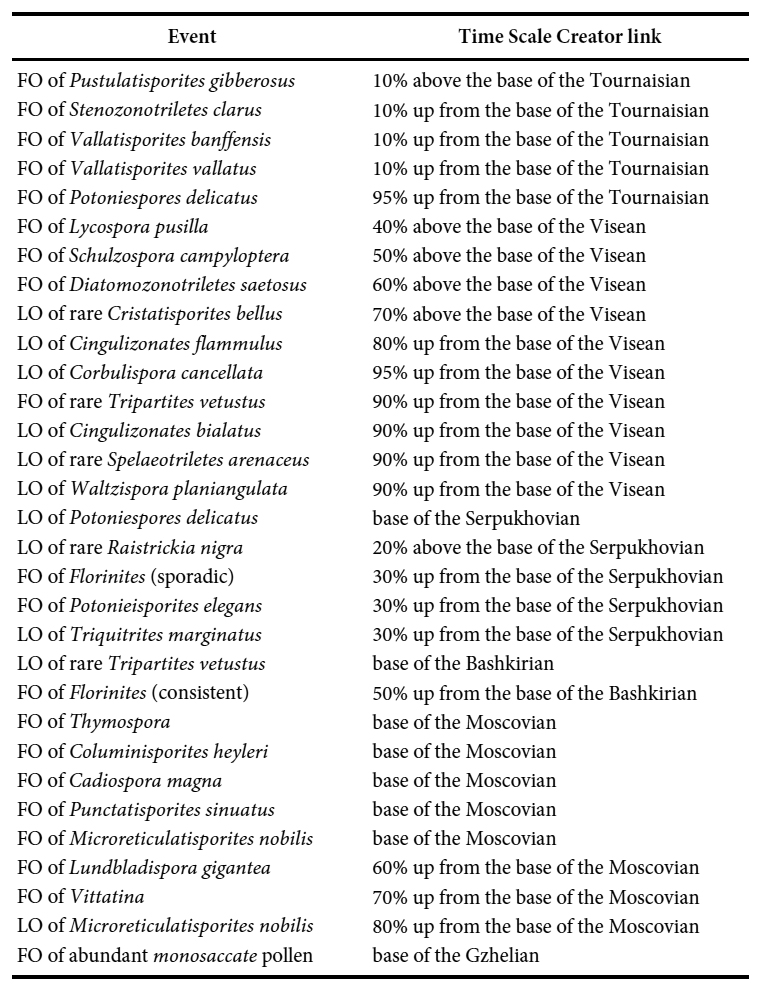
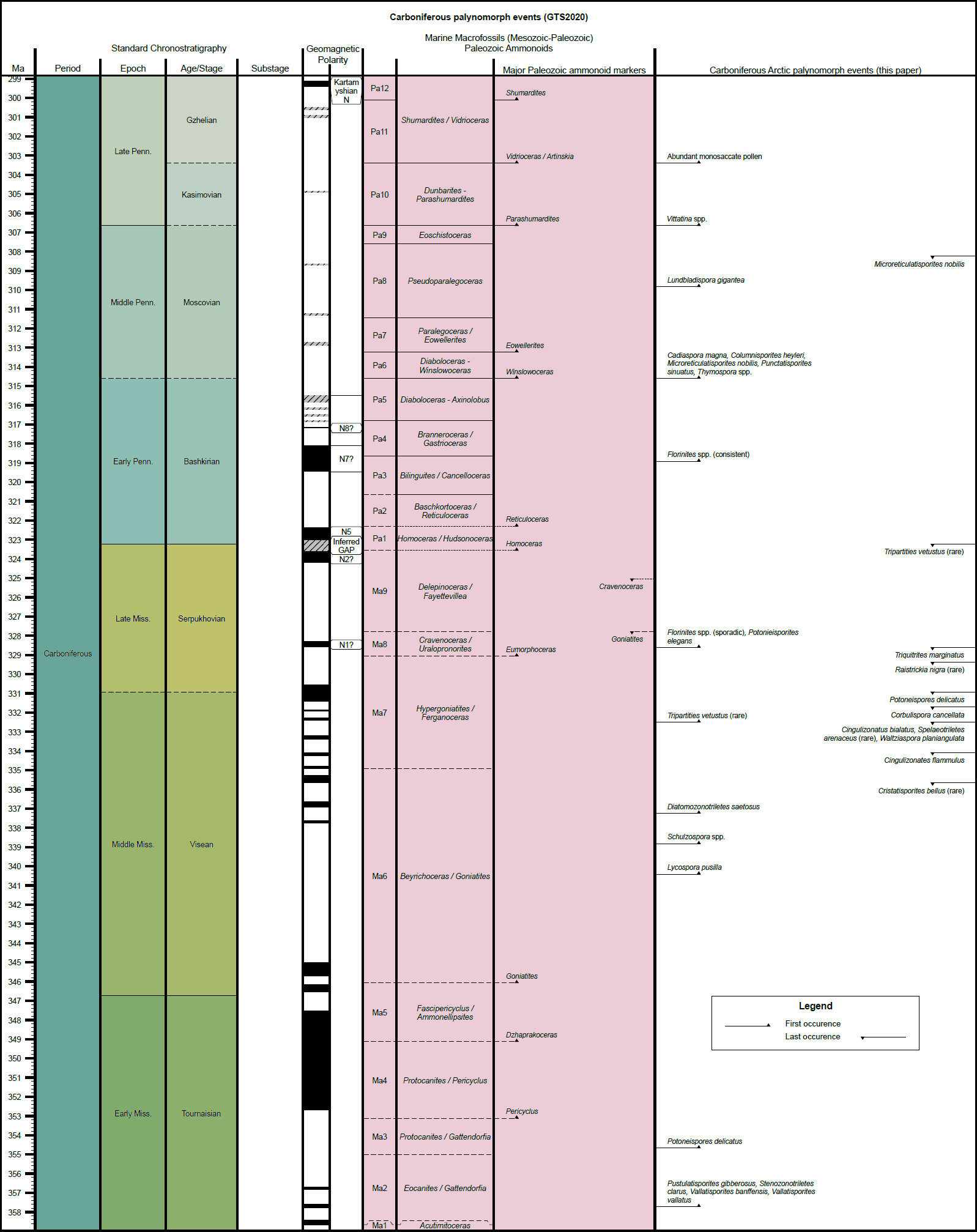
3 The present Carboniferous compilation will be added to others from the CAPE Special Issue of Atlantic Geology, which will contribute ultimately (when all articles in the issue are complete) to the “CAPE datapack” in Time Scale Creator (TSC; https://timescalecreator.org/index/index.php) and thus can be used with other data in TSC make plots such as that shown in Fig. 1. Figure 1 includes the age calibration in millions of years (Ma) according to the 2020 version of TSC.
 Display large image of Figure 1
Display large image of Figure 1
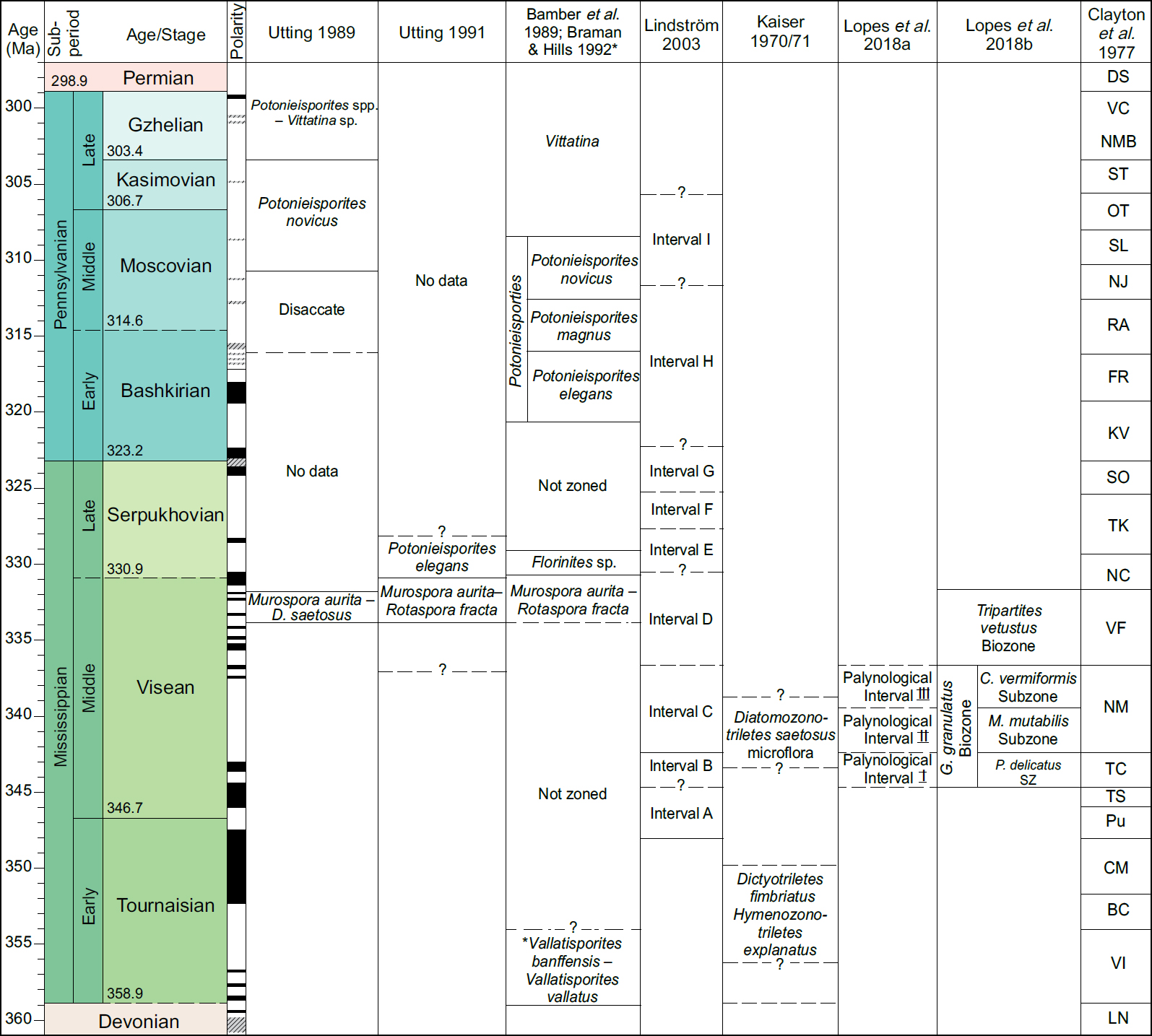
4 The events compiled herein include last occurrences (LOs), first occurrences (FOs), and some abundance events. Where possible, each event is correlated with the base of a chronostratigraphic unit, for example an ammonoid zone or a formal stage. If the event is not equivalent to the base of such a unit, then an estimation is given as a percentage above the base of the chronostratigraphic unit relative to the entire unit. Details of how a biostratigraphic datapack is constructed in TSC from such information are given in Bringué et al. (in press). Localities used for the present compilation are shown in Fig. 2. Figure 3 gives a summary of zonation schemes discussed in the following section and referenced in the Palynoevents section.
 Display large image of Figure 2
Display large image of Figure 2
 Display large image of Figure 3
Display large image of Figure 3
PALYNOSTRATIGRAPHY
5 The events compiled herein include last appearance datums (LOs), first appearance datums (FOs), and some abundance events. Their relationship to other fossil zonal schemes (Fig.2) is shown in the TSC plot (Fig. 3). Despite a re-evaluation by Ogg et al. (2016), the Devonian/Carboniferous boundary is currently formally identified by the first appearance of the conodont Siphonodella sulcata, which appears close to the last worldwide appearance (LO) of the spore Retispora lepidophyta (Paproth et al. 1991; Streel and Marshall 2007; Becker et al. 2012). The LO of Retispora lepidophyta will therefore for practical reasons herein be considered as marking the base of the Carboniferous System. Thus, the lower Tournaisian Tn1 a/b Zone referred to in older papers is considered to be Devonian (e.g., Barss 1967, 1972; Richardson and McGregor 1986) and is not considered in this chapter.
6 In the Sverdrup Basin (Canadian Arctic Islands), palynological research on the Carboniferous began with Utting (1965, 1989) and Utting et al. (1989a, b). Utting (1965) examined material from outcrop samples of the Otto Fiord Formation on Ellesmere Island, which were dated as early Serpukhovian to middle Moscovian based on ammonoids, brachiopods, conodonts and fusulinaceans. The palynomorphs were, however, poorly preserved and their use for palynostratigraphy limited. Utting (1989) established three long-ranging palynozones within the Carboniferous and one spanning the Carboniferous/Permian boundary. These were the Muraspora aurita – Diatomozonotriletes saetosus Assemblage Zone of Visean age, the Disaccate Assemblage Zone of Bashkirian to ?early Moscovian age, the Potonieisporites novicus Assemblage Zone of late Moscovian to possibly Kasimovian age and the Potonieisporites spp. – Vittatina sp. Assemblage Zone of Gzhelian – Asselian of earliest Permian age. The zones were based on outcrop and subsurface samples and were collected from strata in which marine faunas provided some independent age control. However, neither a range chart nor plates were provided. From the Emma Fiord Formation in the Sverdrup Basin, Utting et al. (1989b) recorded well-preserved assemblages and presented range charts, plates and lists of taxa, as well as correlations, and indicated which taxa that they considered to be of stratigraphic importance.
7 Utting (1991) examined 21 outcrops from the Mississippian Hart River Formation of northern Yukon. From these he recorded 32 genera and 44 species. He presented two informal zones: the Murospora aurita – Rotaspora fracta Assemblage Zone of Visean age and the Potonieisporites elegans Assemblage Zone of early Serpukhovian age.
8 From the Northwest Territories (former District of Mackenzie) and Yukon, Braman and Hills (1992) defined the Mississippian Vallatisporites banffensis – Vallatisporites vallatus Biozone from outcrop sections in the Tuttle Formation. They had no independent age control, but the Biozone is above the LO of Retispora lepidophyta, which is close to the Devonian/Carboniferous boundary, so they regarded the Biozone as Tournaisian.
9 From the Alaskan North Slope, Ravn (1991) presented miospores from the Mississippian Kekiktuk Formation derived from conventional cores, sidewall cores and cuttings from 28 wells. He provided an exhaustive systematic section with plates, and described three assemblages from the lower, middle and upper Kekiktuk members respectively. The assemblages were all Visean in age and Ravn compared them to Playford’s (1962, 1963) assemblages from Spitsbergen. He underlined the provincialism of the palynofloras and thus the different ranges in the Arctic compared to those published from other regions.
10 From North-East Greenland Dalhoff et al. (2000) presented two palynological assemblages (Assemblage 1 and 2) from the Mississippian Sortebakker Formation in the Wandel Sea Basin. Earlier, Vigran et al. (1999) had defined palynostratigraphic assemblages covering the uppermost Devonian and Carboniferous from North-East Greenland. Two of these assemblages are of particular interest here as they cover the Tournaisian, an interval relatively poorly represented in the Arctic. These includes the Bascaudaspora spp. Assemblage of late Tournaisian (Tn2-3) age and the Grandispora uncata Assemblage of late Tournaisian (Tn3) age.
11 Pioneering works from the Norwegian Arctic (incorporating the Barents Sea) work include those by Hughes and Playford (1961) and Playford (1962, 1963) from Spitsbergen, and Kaiser (1970, 1971) from Bjørnøya. Later, Lindström (2003) and Bugge et al. (1995) published the first biostratigraphic data from the Carboniferous of offshore Norway.
12 From the Barents Sea area, Bugge et al. (1995) and Lopes et al. (2016, 2018b) based their studies on shallow stratigraphic cores (each up to 580 m long). These cores were drilled with diamond bits, and represent a unique research opportunity for the Norwegian Barents Sea region. All sites tied by high resolution seismic, enabling correlation to conventional seismic (Rise and Sættem 1994). Lopes et al. (2016, 2018a, b) published on the Billefjorden Group both from onshore and offshore areas.
13 Together with a study of macrofloras, Kaiser (1970, 1971) published a comprehensive stratigraphic miospore zonation from Bjørnøya in the Norwegian Arctic, including a systematics section and plates. Playford (1962, 1963) studied 31 palynological samples in three outcrop sections in Spitsbergen (Citadellet, Triungen, and Birger Johnsonfjellet) and presented a systematic section, including plates, comprising 115 species. He defined two palynological assemblages of Visean age, later discussed and incorporated by Lopes et al. (2018a).
14 Lindström (2003) studied a full section of the Billefjorden Group from an exploration well (7120/2-1) on the Loppa High based on conventional core, sidewall core and cuttings samples. She defined nine palynological assemblages (A-I) and correlated her palynological assemblages A-E with the early Viséan to Serpukhovian Pu to TK miospore zones of western Europe. The Pennsylvanian Gipsdalen Group samples from exploration well 7120/2-1 are correlated to the younger SO – NJ zones of Clayton et al. (1977). Scheibner et al. (2012) recorded miospore assemblages from outcrops of the Billefjorden Group in northeastern Spitsbergen, indicating a Famennian and Visean to early Serpukhovian age; however, these were poorly documented as no range-chart, quantitative data or plates are provided.
15 Lopes et al. (2016) defined four assemblages and two subassemblages from the Billefjorden Group on the Finnmark Platform, Barents Sea, based on shallow stratigraphic cores. The assemblages were correlated to the western European biozonation of Clayton et al. (1977) and Clayton (1985) based on taxa abundances and the first and last appearances of selected species. The assemblages are Visean to possibly early Serpukhovian in age. Subsequently, Lopes et al. (2018b) published a revised zonation scheme for the Mississippian succession on the Finnmark Platform, which included new data from exploration wells 7128/4-1 and 7128/6-1.
16 From Spitsbergen Lopes et al. (2018a) presented palynofloras based on 38 samples from the Hørbyebreen and Mumien formations of the Billefjorden Group. The authors integrated work on the same section by Playford (1962).
PALYNOEVENTS
17 A summary of the following events in spreadsheet format is provided as Appendix B herein.
FO of Pustulatisporites gibberosus
18 From studies of outcrop sections of the Tuttle Formation in the Northwest Territories (former District of Mackenzie) and Yukon, Braman and Hills (1992) recorded the FO of Pustulatisporites gibberosus in their youngest Vallatisporites banffensis – Vallatisporites vallatus Assemblage. This record occurs above the LO of Retispora lepidophyta, which marks the Devonian/Carboniferous boundary. Kaiser (1970) reported Pustulatisporites cf. gibberosus from the Visean (“Kulm”) in Bjørnøya. Despite some uncertainty as to its true FO, Pustulatisporites gibberosus seem to have a regional distribution and represents an event with correlation potential.
19 The FO of Pustulatisporites gibberosus is taken to be 10% above the base of the Tournaisian.
FO of Stenozonotriletes clarus
20 From studies of outcrop sections of the Tuttle Formation in the Northwest Territories (former District of Mackenzie) and Yukon, Braman and Hills (1992) recorded the FO of Stenozonotriletes clarus in their youngest Vallatisporites banffensis – Vallatisporites vallatus Assemblage. This record is above the FO of Retispora lepidophyta marking the Devonian/Carboniferous boundary. Stenozonotriletes clarus is seen in younger deposits in the Barents Sea area, but its FO is not recorded from this area.
21 The FO of Stenozonotriletes clarus is taken as 10% up from the base of the Tournaisian.
FO of Vallatisporites banffensis
22 From studies of outcrop sections of the Tuttle Formation in the Northwest Territories (former District of Mackenzie) and Yukon, Braman and Hills (1992) recorded the FO of Vallatisporites banffensis from their youngest Vallatisporites banffensis – Vallatisporites vallatus Assemblage. This record is above the LO of Retispora lepidophyta marking the Devonian/Carboniferous boundary. This taxon, whose regional distribution was discussed by Playford and Melo (2012), seems to have its earliest occurrence close to the Devonian/ Carboniferous boundary in northernmost North America.
23 The FO of Vallatisporites banffensis is taken as 10% up from the base of the Tournaisian.
FO of Vallatisporites vallatus
24 From studies of outcrop sections of the Tuttle Formation in the Northwest Territories (former District of Mackenzie) and Yukon, Braman and Hills (1992) recorded the FO of Vallatisporites vallatus from their youngest Vallatisporites banffensis – Vallatisporites vallatus Assemblage. This record is above the LO of Retispora lepidophyta marking the Devonian/Carboniferous boundary. This taxon, whose regional distribution was discussed by Playford and Melo (2012), seems to have its earliest occurrence close to the Devonian/Carboniferous boundary in northernmost North America.
25 The FO of Vallatisporites vallatus is taken as 10% up from the base of the Tournaisian.
FO of Potoniespores delicatus
26 The FO of Potoniespores delicatus is recorded from several locations in the Arctic. In Spitsbergen, Lopes et al. (2018a) recorded this species from the middle part of the Hørbyebreen Formation, corresponding to the lower part of the NM Zone of the western European zonation. In the Barents Sea area, Lopes et al. (2016, 2018b) recorded Potoniespores delicatus in the Soldogg Formation. Ravn (1991) recorded it from the basal Kekiktuk Formation in Alaska, which is close to the Tournaisian/Visean boundary.
27 The FO of Potoniespores delicatus is taken as 95% up from the base of the Tournaisian.
FO of Lycospora pusilla
28 From the Sverdrup Basin, Utting (1989) reported that Lycospora pusilla partly dominated assemblages from the Emma Fiord Formation. Utting et al. (1989b) also recorded Lycospora pusilla from the base of the Emma Fiord Formation, but the base is above an unconformity; so the true base of the species is uncertain. In Spitsbergen, Lycospora pusilla first appears within the middle Visean (Lopes et al. 2018a), correlating with the TC biozone. From the Alaskan North Slope, Ravn (1991) recorded Lycospora pusilla from the middle part of the lower Kekiktuk Formation. Lycospora pusilla was also recorded from the Visean in eastern North Greenland (Dalhoff et al. 2000).
29 The FO of Lycospora pusilla is taken as 40% above the base of the Visean.
FO of Schulzospora campyloptera
30 In Spitsbergen, Lopes et al. (2018a) recorded Schulzospora campyloptera from the upper part of the Høybyebreen Forma-tion, corresponding to the middle part of the NM biozone of western Europe. From the Finnmark Platform, Lopes et al. (2016) recorded Schulzospora in assemblages correlated to the TC biozone. Lindström (2003) reported Schulzospo-ra spp. from ditch cuttings that she dated to the early/late Visean (based on correlation with the TC biozone in the western European Zonation). Schulzospora campyloptera is also recorded in eastern North Greenland from the Visean (Dalhoff et al. 2000) and correlated with the middle part of the NM Biozone.
31 The FO of Schulzospora campyloptera is taken as 50% above the base of the Visean.
FO of Diatomozonotriletes saetosus
32 Utting (1989) reported Diatomozonotriletes saetosus from the middle part of the Emma Fiord Formation in the Sverdrup Basin, whereas Utting et al. (1989b) recorded it almost from the base of the Emma Fiord Formation. Lopes et al. (2016) recorded Diatomozonotriletes saetosus in the Tettegras Formation, correlated to the Middle/Upper Visean. In the Høybyebreen Formation on Spitsbergen, Diatomozonotriletes saetosus correlated to the NM Biozone (Lopes et al. 2018a). Lindström (2003) recorded the taxon in her Assemblage G, whereas in eastern North Greenland, Dalhoff et al. (2000) recorded it from the upper part of the Sorterbakker Formation, which correlates to the NM Zone of the western European Biozonal scheme of Clayton et al. (1977). Ravn (1991) recorded it from the middle part of the Kekiktuk Formation.
33 The FO of Diatomozonotriletes saetosus is taken as 60% above the base of the Visean.
LO of rare Cristatisporites bellus
34 In the Barents Sea area, rare Cristatisporites bellus is recorded from several locations. Kaiser (1970) recorded the species in his Visean 3 in Bjørnøya, approximately corresponding to the TC–VF zones. Lopes et al. (2016, 2018a, b) recorded Cristatisporites bellus extending to the base of the Blærerot Formation in exploration well 7128/6-1, and in the middle of the Hørbyebreen Formation.
35 The LO of rare Cristatisporites bellus is taken as 70% above the base of the Visean.
LO of Cingulizonates flammulus
36 Ravn (1991) described this taxon as extending to the middle part of the Kekiktuk Formation of Alaska, based on material from cuttings. In the Barents Sea area, common Cingulizonates flammulus was recorded from shallow core material (Lopes et al. 2016) and cuttings (Lopes et al. 2018b) extending to the base of the Blærerot Formation in exploration well 7128/6-1. Lopes at al. (2018a) also recorded this species from their interval III in outcrop sections in Spitsbergen, corresponding to the NM Zone from the Mumien Formation (Birger Johnsonfjellet member).
37 The LO of Cingulizonates flammulus is taken as 80% up from the base of the Visean.
LO of Corbulispora cancellata
38 This taxon was recorded by Ravn (1991) as spanning the Kekiktuk Formation on the Alaskan North Slope. Utting et al. (1989b) reported it (as Reticulatisporites cancellatus) spanning the Emma Fiord Formation. In Spitsbergen, Lopes et al. (2018a) recorded Corbulispora cancellata spanning the Hørbyebreen and Mumien formations. In the Barents Sea, Lopes et al. (2016) recorded this species as extending just up to the base of the Blærerot Formation from shallow core 7129/3-U-1. It was also recorded in cuttings samples from exploration well 7128/6-1, in an interval correlated to the NM Zone of western Europe.
39 The LO of Corbulispora cancellata is taken as 95% up from the base of the Visean.
FO of rare Tripartites vetustus
40 The FO of rare Tripartites vetustus is based on Clayton et al. (1990), who recorded six Mississippian palynological events that can be correlated around the Northern Hemisphere. One was the FO of Tripartites vetustus, which they reported to be intra-upper Visean, at the top of the VF Zone. Tripartites vetustus has so far not been reported from the Sverdrup Basin, and seems to be rare in the Barents Sea. The occurrence of this species was recorded by Lopes et al. (2016, 2018a) from shallow core 7029/3-U-1, as well as from exploration wells in the lower part of the Blærerot Formation on the Finnmark Platform.
41 The FO of rare Tripartities vetustus is taken as 90% up from the base of the Visean.
LO of Cingulizonates bialatus
42 Utting (1989) recorded this species from several sections in the Emma Fjord Formation in the Sverdrup Basin. The species is also listed as important in Utting’s (1989) Muraspora aurita – Diatomozonotriletes saetosus Assemblage Zone, correlated to the older NM Zone or possibly the VF Zone. Utting refers to correlations with marine vertebrate faunas in beds of a late Visean age. Bamber et al. (1989) also listed this species as stratigraphically important in their Murospora aurita – Rotaspora fracta Assemblage Zone, to which they assign a late Visean (Visean 3) age. Lindström (2003, p. 336) recorded this species to the top of her Interval E, in the upper part of the Billefjorden Group, to which she assigned an age “no younger than the TK Miospore Zone”. In Spitsbergen, the LO of Cingulizonates bialatus is in the upper part of the Mumien Formation, corresponding to the NM Zone, although because of an overlying hiatus this does not reflect the true last occurrence. Lopes et al. (2018a) recorded Cingulizonates bialatus ranging up to within the Blærerot Formation in wells from the Finnmark Platform. They correlated their assemblage to the VF Miospore Zone of western Europe. Ravn (1991) recorded rare specimens extending into the upper part of the Kekiktuk Formation in Alaska. In East Greenland in extends into the upper part of the Sorterbakker Formation (Dalhoff et al. 2000).
43 The LO of Cingulizonates bialatus is taken as 90% up from the base of the Visean.
LO of rare Spelaeotriletes arenaceus
44 Utting (1989) and Utting et al. (1989b) recorded Spelaeotriletes arenaceus in several outcrop sections from the Emma Fiord Formation in the Sverdrup Basin. It is listed as important in Utting’s (1989) Muraspora aurita – Diatomozonotriletes saetosus Assemblage Zone, which correlates to the older NM Zone or possibly the VF Zone. Utting referred to correlations with marine vertebrate faunas in beds of a late Visean age. Applying the approach described by Playford et al. (2001) to identify this species, Lopes et al. (2018a) recorded it ranging up into the Blærerot Formation in wells from the Finnmark Platform. They correlated their assemblage to the VF Miospore Zone of western Europe.
45 The LO of rare Spelaeotriletes arenaceus is taken as 90% up from the base of the Visean.
LO of Waltzispora planiangulata
46 Utting et al. (1989b) recorded this species in several sections from the Emma Fjord Formation in the Sverdrup Basin. The species as also listed as important in Utting’s (1989) Muraspora aurita – Diatomozonotriletes saetosus Assemblage Zone, which correlates to the older NM Zone or possibly the VF Zone. He refers to correlations with marine vertebrate faunas in beds of late Visean age. Lopes et al. (2018b) recorded this species ranging up into the Blærerot Formation in wells from the Finnmark Platform. They correlated their assemblage to the VF Miospore Zone of western Europe. Kaiser (1970) also recorded this species from the Visean (“Kulm”) of Bjørnøya. Dalhoff et al. (2000) recorded Waltzispora planiangulata in the Sorterbakker Formation in eastern North Greenland.
47 The LO of Waltzispora planiangulata is taken as 90% up from the base of the Visean.
LO of Potoniespores delicatus
48 The LO of Potoniespores delicatus is recorded from several locations in the Arctic. In Spitsbergen, Lopes et al. (2018a) recorded rare Potoniespores delicatus up to the middle to upper part of the Mumian Formation, corresponding to the NM Zone of the western European zonation. In the Barents Sea area, Lopes et al. (2016, 2018b) recorded Potoniespores delicatus up into the upper part of the Tettegras Formation, corresponding to the NM Zone of western Europe. From Bjørnøya, Kaiser (1970) recorded it from the late Visean (“Kulm”). Lindström (2003) listed an occurrence of Potonie-spores delicatus in the Gipsdalen Group but, as it was based on one cutting sample and one side wall core, it could repre-sent reworking and therefore its LO there is uncertain.
49 The LO of Potoniespores delicatus is taken as the base of the Serpukhovian.
LO of rare Raistrickia nigra
50 Rare specimens of Raistrickia nigra are recorded from various locations in the Arctic. Lopes et al. (2018a) recorded this species ranging up to somewhere within the Blærerot Formation in wells from the Finnmark Platform, Barents Sea; this corresponds to just above the VF Zone of Clayton et al. (1977). Lindström (2003) recorded the species in her Assemblage G, which is Serpukhovian. From the Wandel Sea Basin in eastern North Greenland, Dalhoff et al. (2000) recorded Raistrickia nigra from one sample in the Sortebakker Formation, which they correlated to the NM Zone of Clayton et al. (1977). Raistrickia nigra has not yet been reported from the Canadian Arctic. Whether the LO is a cir-cum-Arctic event is therefore uncertain.
51 The LO of rare Raistrickia nigra is taken as 20% above the base of the Serpukhovian.
FO of Florinites (sporadic)
52 The FO of sporadic Florinites spp. is based on Bamber et al. (1989), who reported the FO of Florinites sp. in their Florinites sp. Assemblage Zone of early Serpukhovian age from the uppermost Hart River Formation in northern Yukon. Clayton et al. (1977) recorded Florinites spp. from the middle of their NC Miospore Zone of western Europe, which they correlated to the Namurian (within the Serpukhovian). According to McLean et al. (2005), Florinites has its FO around the base of the late Serpukhovian (within the late Arnsbergian).
53 The FO of sporadic Florinites spp. is taken as 30% up from the base of the Serpukhovian.
FO of Potonieisporites elegans
54 Utting (1991) described the Potonieisporites elegans Assemblage Zone from the upper part of the Hart River Formation in northern Yukon. He discussed the problems with correlation, but tentatively correlated the Assemblage Zone to the NC Zone of Clayton et al. (1977), pointing out the lack of many of the characteristic species from the latter. Lindström (2003) referred to records of Potonieisporites elegans from the Efuglvika Member of the Kapp Kåre Formation on Bjørnøya based on personal communication from J.O.Vigran.
55 The FO of Potonieisporites elegans is taken as 30% up from the base of the Serpukhovian.
LO of Triquitrites marginatus
56 Lopes et al. (2018b) reported the youngest occurrence of Triquitrites marginatus from two exploration wells penetrating the Blærerot Formation of the Billefjorden Group on the Finnmark Platform. It appears in their Monilospora mutabilis Subzone of late Visean age, which correlates to the lower part of the NM Zone of the western European zonation (Clayton et al. 1977). Lopes et al. (2018b) stated that Triquitrites marginatus belongs to a group of taxa, each of which seems to have a similar stratigraphic range in both the Finnmark Platform and in western Europe. Lindström (2003) reported the FO of Triquitrites marginatus from the top of her Interval E, which corresponds to the top of the Billefjorden Group. She assigned this interval to the early Serpukhovian based on correlation to the Clayton et al. (1977) TK Miospore Zone of western Europe.
57 The LO of Triquitrites marginatus is taken as 30% up from the base of the Serpukhovian.
LO of rare Tripartites vetustus
58 Tripartites vetustus is recorded in low numbers from the lowermost part of the Blærerot Formation on the Finnmark Platform (Lopes et al. 2018b), but the species has so far not been reported from the Sverdrup Basin. Clayton et al. (1990) published six palynological events in the Mississippian that can be correlated around the Northern Hemisphere. One of these is the extinction of Tripartites vetustus, which they reported as within the lower Namurian (Serpukhovian).
59 The LO of rare Tripartites vetustus is taken as the base of the Bashkirian.
FO of Florinites (consistent)
60 The first consistent occurrences of Florinites spp. seem to be within the Bashkirian. Bamber et al. (1989) reported abundant monosaccate pollen, including Florinites spp., from their Potonieisporites Assemblages Zone of Bashkirian age in northern Yukon – northwestern Northwest Territories (former District of Mackenzie). The base of consistent to common Florinites spp. is recorded by Lindström (2003) in her “Interval I” from the Loppa High (exploration well 7120/2-1) near the top of the Gipsdalen Group. According to the western European miospores zonation of Clayton et al. (1977), the consistent occurrence of Florinites spp. occurs at the base of the RA Miospore Zone (Clayton et al. 1977). From North-East Greenland, Vigran et al. (1999) reported Florinites from their Potonieisporites spp., Radiizonates aligerens – Schulzospora rara, Savitrisporites Concurrent Range Zone, which may correlate to the lower part of the RA Miospore Zone of Clayton et al. (1977). However this correlation is problematic (see discussion in Vigran et al. 1999).
61 The FO of consistent Florinites spp. is taken as 50% up from the base of the Bashkirian.
FO of Thymospora
62 Bamber et al. (1989) listed the monolete spore genus Thymospora as having a base in their Potonieisporites magnus Assemblage Subzone of late Bashkirian to Moscovian age. The base of Potonieisporites magnus was recorded by Lindström (2003) in her “Interval I” from the Barents Sea (exploration well 7120/2-1), near the top of the Gipsdalen Group. Her correlation is based on palynology, and the FO is correlated to the base of the NJ Miospore Zone in the western European scheme of Clayton et al. (1977).
63 The FO of Thymospora is taken as the base of the Moscovian.
FO of Columinisporites heyleri
64 From the Barents Sea (exploration well 7120/2-1), Lindström (2003) recorded Columnisporites heyleri from core material in her “Interval I” near the top of the Gipsdalen Group. She correlated it to the base of the NJ Miospore Zone of western Europe (Clayton et al. 1977). No older records are reported from the Arctic.
65 The FO of Columinisporites heyleri is taken as the base Moscovian.
FO of Cadiospora magna
66 Lindström (2003) recorded Cadiospora magna in her “Interval I” near the top of the Gipsdalen Group from conventional core material in exploration well 7120/2-1 from the Barents Sea. She correlated the base of “Interval I” to the base of the NJ Miospore Zone of western Europe (Clayton et al. 1977). No older records have been reported from the Arctic.
67 The FO of Cadiospora magna is taken as the base of the Moscovian.
FO of Punctatisporites sinuatus
68 From conventional core in Barents Sea exploration well 7120/2-1, Lindström (2003) recorded Punctatisporites sinuatus from her “Interval I”, near the top of the Gipsdalen Group. Its FO correlates with the base of the NJ Miospore Zone of western Europe (Clayton et al. 1977). No older records have been reported from the Arctic.
69 The FO of Punctatisporites sinuatus is taken as the base of the Moscovian.
FO of Microreticulatisporites nobilis
70 From core in exploration well 7120/2-1 in the Barents Sea, Lindström (2003) recorded Microreticulatisporites nobilis from her “Interval I” near the top of the Gipsdalen Group. Her correlation, based on palynology, is to the base of the NJ Miospore Zone of western Europe (Clayton et al. 1977). No older records have been reported from the Arctic.
71 The FO of Microreticulatisporites nobilis is taken as the base of the Moscovian.
FO of Lundbladispora gigantea
72 From core material in Barents Sea exploration well 7120/2-1, Lindström (2003) recorded Lundbladispora gigantea “her Interval I” near the top of the Gipsdalen Group. She correlated this occurrence with the SL Miospore Zone of western Europe (Clayton et al. 1977), indicating a late Westphalian to Stephanian age. No older records have been reported from the Arctic.
73 The FO of Lundbladispora gigantea is taken as 60% up from the base of the Moscovian.
FO of Vittatina
74 Utting (1989) recorded common Vittatina from the Middle Canyon Fiord Formation. Conodonts suggested a Gzhelian to Asselian age for this formation (Utting 1989, p. 238). Bamber et al. (1989) recorded their Vittatina Assemblage Zone from the late Moscovian of northern Yukon. The FO of Vittatina was also recorded below the Tyrellfjellet Member in the Nordenskiöldbreen Formation from Nordaustlandet, Svalbard by Mangerud and Konieczny (1991).
75 The FO of common Vittatina is taken as 70% up from the base of the Moscovian.
LO of Microreticulatisporites nobilis
76 Microreticulatisporites nobilis was recorded by Lindström (2003) in her “Interval I”, near the top of the Gipsdalen Group from Barents Sea exploration well 7120/2-1. Based on palynology, the youngest occurrence of Microreticulatisporites nobilis was calibrated to occur near the top of the SL Miospore Zone of western Europe (Clayton et al. 1977). Since this is the only record of this species from the Arctic so far, the LO is uncertain.
The LO of Microreticulatisporites nobilis is here taken as 80% up from the base of the Moscovian.
FO of abundant monosaccate pollen
77 Data from various areas in the present Arctic indicate that the FO of abundant monosaccate pollen occurs in the latest Carboniferous. It is recorded from the uppermost Canyon Fiord Formation, where conodonts indicate a Gzhelian to Asselian age (A.C. Higgins, pers. comm. 1986 in Utting 1989). However, the age of the FO is uncertain as independent data is sparse. Similar pollen and spore assemblages in the upper part of the unnamed equivalent of the Ettrain Formation (southern Eagle Plain) and the lower part of the Jungle Creek Formation (northern Ogilvie Mountains) of northern Yukon (Bamber et al. 1989) were assigned a Gzhelian age based on brachiopods (Waterhouse and Waddington 1982). Consistent monosaccate pollen were also recorded from the Idunfjellet member of the Nordenskiöldbreen Formation, Nordaustlandet, Svalbard, from a similar age (Mangerud and Konieczny 1991). Lindström (2003) reported assemblages with 30% monosaccate pollen in the upper part of her assemblage I. The age assignment of this youngest assemblage is uncertain, although she tentatively inferred a possible Moscovian or Kasimovian–Gzhelian age.
78 The FO of abundant monosaccate pollen is taken as the base of the Gzhelian.
ACKNOWLEDGEMENTS
We are grateful to Eva Bjørseth (University of Bergen) and Bill MacMillan (Geological Survey of Canada ― Atlantic, who helped draft the figures. We thank Geoff Playford and Geoff Clayton for their constructive reviews; and Manuel Bringué for help with the TSC plot (Fig. 1). This article is a contribution to the TransGEM event stratigraphy activity of the Geological Survey of Canada’s Geo-mapping for Energy and Minerals Program.
REFERENCES
Editorial responsibility: Robert A. Fensome