Articles
Filicopsida from the lower Westphalian (Middle Pennsylvanian) of Nova Scotia and New Brunswick, Maritime Provinces, Canada
doi:10.4138/atlgeol.2019.001
ABSTRACT
This paper presents a taxonomic revision of filicopsid taxa from the lower to middle Westphalian strata of Nova Scotia and New Brunswick, Maritime Provinces of Canada. Most of the material represents sporadic historical collecting by Geological Survey of Canada (GSC) personnel, and specimens are in the GSC collections in Ottawa. Additional specimens are in the New Brunswick Museum at Saint John, the Fundy Geological Museum at Parrsboro, and the Joggins Fossil Institute at Joggins. Two specimens from outside Canada clarify specific characteristics. The revision involved the detailed examination of 20 adpression (mainly impression) taxa, of which one, Germera brousmicheae, is new. Detailed synonymy lists, with particular focus on records from Canada and the USA, facilitate a refinement of the stratigraphic and geographic distribution of these species. Most of the taxa from the Maritimes are the same as those from other parts of the paleoequatorial belt of Pennsylvanian times. The Maritimes record of filicopsid taxa closely resembles the filicopsid floras of western Europe, most notably the British Isles.
RÉSUMÉ
Cet article présente une révision taxonomique des taxons de Filicopsida provenant de la strate westphalienne moyenne à inférieure de la Nouvelle-Écosse et du Nouveau-Brunswick, des provinces maritimes du Canada. La plupart des documents décrivent la collecte de données historiques sporadiques effectuée par les membres du personnel de la Commission géologique du Canada (CGC), et des spécimens se trouvent dans les collections de la CGC à Ottawa. D’autres spécimens sont exposés au Musée du Nouveau-Brunswick de Saint John, au Fundy Geological Museum de Parrsboro, et au Joggins Fossil Institute de Joggins. Deux spécimens prélevés à l’extérieur du Canada apportent des précisions sur des caractéristiques particulières. La révision a nécessité l’examen de 20 taxons en adpression (principalement en impression), parmi lesquels un taxon, Germera brousmicheae, est nouveau. Des listes détaillées de synonymes, mettant un accent particulier sur les registres provenant du Canada et des États-Unis, facilitent le raffinement de la répartition géographique et stratigraphique de ces espèces. La plupart des taxons des Maritimes sont les mêmes que ceux des autres parties de la ceinture paléoéquatoriale du Pennsylvanien. Les registres de taxons de Filicopsida provenant des Maritimes ressemblent fortement aux flores de la classe des Filicopsida de l’Europe de l’Ouest, plus particulièrement des îles Britanniques.
[Traduit par la redaction]
INTRODUCTION
1 This study is the eighth part of a taxonomic revision of the upper Namurian to middle Westphalian floras of the Maritime Provinces of Canada. The revision was embarked upon in 2000 on the initiative of the late John Utting, then of the Geological Survey of Canada. The reporting of results began with a series of short papers (Wagner 2001a, 2005a, b, 2008; Wagner and Álvarez-Vázquez 2008), and continued with more extensive contributions on lycopsids (Álvarez-Vázquez and Wagner 2014) and the sphenopsid genera Annularia and Asterophyllites (Álvarez-Vázquez and Wagner 2017). (All names of taxa at generic and lower rank cited herein are listed with full authorship in the Appendix).
2 The floral remains dealt with in this study are preserved as adpressions (mainly impressions). Most of the material recorded here corresponds to that recorded by Walter A. Bell in the early part of the twentieth century (Bell 1944, 1966), with specimens reposited in the collections of the Geological Survey of Canada, Ottawa. Also studied were specimens from the New Brunswick Museum at Saint John, the Fundy Geological Museum at Parrsboro, Nova Scotia, and the Donald Reid collection in the Joggins Fossil Institute, Joggins, Nova Scotia.
3 Although Bell (1944) published a very comprehensive account, his descriptions were brief and illustrations, although numerous and at natural size, do not always provide the necessary detail for an incontrovertible identification. The paper by Bell (1966) is more useful because the specimens illustrated were selected for quality. Much more information has become available on late Namurian to middle Westphalian plant macrofossils in recent decades, making Bell’s work of seven decades ago in need of revision.
REPOSITORY OF SPECIMENS, LOCALITY AND CATALOGUE NUMBERS
4 Most of the specimens reviewed in the present paper are in the collections of the Geological Survey of Canada, Ottawa (catalogue numbers preceded by “GSC”). For more complete information about the localities the reader is referred to Bell (1938, p. 108–115; 1944, p. 111–118; 1962, p. 63–64). Additional material from the collections of the New Brunswick Museum at Saint John has catalogue numbers preceded by “NBMG”. Moreover, six specimens from the Fundy Geological Museum, Parrsboro, Nova Scotia (“FGM”) have been studied, as well as four from the Donald Reid Collection (“DRC”), Joggins Fossil Institute, Joggins, Nova Scotia.
SYSTEMATICS
5 As in our previous revisions of lycopsids (Álvarez-Vázquez and Wagner 2014) and Annularia and Asterophyllites (Álvarez-Vázquez and Wagner 2017), selective lists of synonyms are provided with the emphasis on published records from Canada and the USA. Only the most significant records from outside Canadian and the USA and/or specimens in the possession of the present author are cited, as well as synonyms accepted by the present author. The reader is referred to the Fossilium Catalogus (Plantae) for more complete (up to a certain date), but uncritical records.
6 Annotations in the synonymy lists are as follows: * = protologue; § = first publication of currently accepted combination; T = other illustrations of the holotype; ? = references to doubtful specimens due to poor illustration or preservation; p = pars — only part of the specimens published belong to the species; v = vide — the author has seen the specimen(s); k = reference includes cuticular evidence; acc. = according.
7 Descriptions, comparisons and remarks on published and revised specimens are given, as well as the stratigraphic and geographic distribution of taxa and their occurrences in Canada and the USA. Stratigraphic attributions are in accordance with the western European regional chronostratigraphic scheme of the Pennsylvanian Subsystem, since this is the most relevant to fossil plant records.
Class Filicopsida
Order Crossothecales Danzé 1956
Family Crossothecaceae Danzé 1956
Genus Crossotheca Zeiller 1883
- 1883 Crossotheca Zeiller, p. 180 (August).
- 1883 Sorotheca Stur, p. 175 [807] (December).
- 1956 Crossotheca Zeiller; Danzé, p. 55–57, 135–139.
- 1982 Crossotheca Zeiller; Brousmiche, p. 686–688.
- 1983 Crossotheca Zeiller; Brousmiche, p. 120–122.
- 1986 Crossotheca Zeiller; Brousmiche, p. 233–234.
8 Type.Crossotheca crepinii Zeiller 1883.
9 Remarks. Pedunculate eusporangiate synangial fertile structures that are of very variable appearance depending on the degree of maturity: they may be globular, spherical or elongate. Individual lamina consists of a thick, horizontal, narrow part and a large folded part with segments that protect the sporangia; these are long and inserted in small groups of four or five terminal to veins. Vegetative pinnules sphenopteroid or pecopteroid.
Crossotheca crepinii Zeiller 1883
(Figs. 1a–b, d)
 Display large image of Figure 1
Display large image of Figure 1
- * 1883 Crossotheca Crepini Zeiller, p. 181, pl. IX, figs. 1–9.
- T 1886–88 Sphenopteris (Crossotheca) Crepini Zeiller, p. 112–115, pl. XIII, figs. 1–1D (lectotype as designated by Brousmiche 1983), figs. 2–3A.
- * 1886–88 Sphenopteris (Crossotheca) Boulayi Zeiller, p. 115–117, pl. IV, figs. 4–4C (acc. to Brousmiche 1983).
- 1938 Crossotheca boulayi Zeiller; Bell, p. 47, pl. XXXVI, figs. 1–4 (acc. to Brousmiche 1982).
- 1938 Corynepteris winslovii White; Bell, p. 39, pl. XXV, figs. 1–4.
- * 1938 Sphenopteris aculeata Bell, p. 29, pl. XI, fig. 5; pl. XII, fig. 1 (although difficult to judge from the illustration, Bell’s description suggests that this species falls within the morphological variation of Crossotheca crepinii. Additional specimens figured by Bell in 1966 from the same area in the Sydney coalfield confirm this impression).
- * v 1944 Sphenopteris philipensis Bell, p. 64–65, pl. VIII, fig. 2; pl. IX (partly refigured here as Fig. 1a); pl. X, figs. 2, 4 (fragmentary); pl. XII, fig. 3 (Fig. 1d herein).
- T 1956 Crossotheca Crepini Zeiller; Danzé, p. 139–146, pl. I, figs. 1–5b; pl. II, fig. 1 (same as Zeiller 1886, pl. XIII, fig. 1), fig. 2 (same as Zeiller 1883, pl. IX, fig. 1), figs. 3–3a; pl. III, figs. 1–1a, fig. 2 (same as Corsin 1932, pl. 30, fig. 2), figs. 3–5a; pl. IV, figs. 1–2; pl. V, figs. 1–4b; text-figs. 2a–d.
- 1956 Crossotheca Boulayi Zeiller; Danzé, p. 151– 157, pls VII, figs. 1–1d; pl. VIII, figs. 1–6a; pl. IX, figs. 1–2a; pl. X, figs. 1–4a; text-figs. 4a–d.
- * 1956 Crossotheca Bourozi Danzé, p. 157–160, pls XI, figs. 1–6a; pl. XII, figs. 1–4b; pl. XIII, figs. 1–2c; pl. XIV, figs. 1–3; pl. XV, figs. 1–2a; text-figs. 5a–c (acc. to Brousmiche 1983).
- 1966 Sphenopteris (Crossotheca) boulayi Zeiller; Bell, p. 42, pl. XX, figs. 7–8.
- 1966 Sphenopteris aculeata Bell, p. 52, pl. XXV, figs. 2–3.
- ? 1981 Sphenopteris (?) schatzlarensis (Stur) Zeiller; DiMichele and Dolph, p. 5, pl. 1, fig. 7 (difficult to judge from the illustration).
- T 1982 Crossotheca crepinii Zeiller; Brousmiche, p. 688–691 (including synonymy), pl. 1, fig. 1 (same as Danzé 1956, pl. VII, figs. 1a–1d), figs. 2–2a (same as Danzé 1956, pl. VIII, figs. 3–3a), figs. 3–3a (same as Zeiller 1886, pl. IV, fig. 4), figs. 4–4a (same as Danzé 1956, pl. I, fig. 1), fig. 5 (same as Zeiller 1886, pl. XIII, figs. 1–1b), figs. 6–6a (same as Danzé 1956, pl. I, fig. 4); pl. 2, figs. 1–1b (same as Danzé 1956, pl. XIII, figs. 2–2c), figs. 2–4a, figs. 5–5a (same as Danzé 1956, pl. XIV, fig. 3), figs. 6–6a; pl. 3, fig. 1 (same as Danzé 1956, pl. V, figs. 4–4b); pl. 2, fig. 2 (same as Danzé 1956, pl. IV, figs. 1–1a), fig. 3 (same as Danzé 1956, pl. IX, figs. 2–2a), fig. 4 (same as Zeiller 1886, pl. IV, fig. 4), figs. 5–7; pl. 4, figs. 1–2b, figs. 3–8 (spores); pl. 5, figs. 1–8 (spores); text-figs. 1A–3.
- T 1983 Crossotheca crepinii Zeiller; Brousmiche, p. 122–126 (including synonymy), pl. 24, figs. 1–1a (part of the lectotype), figs. 2–2a (same as Zeiller 1886, pl. IV, fig. 4), figs. 3–3a, figs. 4–4a (same as Danzé 1956, pl. VIII, figs. 1–1a), figs. 5–8a; pl. 25, figs. 1–4a, 5 (enlargement of Zeiller 1886, pl. IV, fig. 4), figs. 6–8a; pl. 26, figs. 1–1g; text-figs. 30A–J3.
10 Description. Frond at least tripinnate. Antepenultimate rachis straight, flat, ca. 4 mm wide, finely striate longitudinally and with faint transverse markings. Penultimate pinnae well-spaced, alternate, always incomplete but apparently lanceolate. Dimensions: at least 60 mm long and 30 mm broad at base. Penultimate rachis inserted at 45–50°, straight, flat, ca. 1 mm wide, with fine longitudinal striations and broad transverse markings (not always visible). Last order pinnae closely spaced but not touching laterally, lanceolate, with a rounded apical pinnule; up to 16 mm long and 7 mm broad. Last order rachis inserted at 70–80°, straight, flat, ca. 0.6 mm wide. Pinnules alternate and slightly oblique to the rachis; small, with a stout footstalk, and apparently linked by a narrow strip of lamina alongside the rachis; pinnules deeply dissected into three to nine rounded to spatulate lobes. Nervation hardly visible due to the thick convex lamina and the coarseness of the siltstone on which the pinnules are imprinted. When evident, the midrib is relatively wide, slightly decurrent, forking once or twice at a wide angle.
11 Remarks. Bell (1944) figured one small fragment and two larger ones, the latter consisting of poorly preserved vegetative remains of a species that he regarded as new, i.e. Sphenopteris philipensis. Bell’s specimens are impressions on a siltstone that emphasizes the convex nature of pinnule lobes. The relative coarseness of sediment and the vaulted lamina combine to show the exact outline of pinnules only occasionally. Unfortunately the specimens were varnished, thus making detailed observation as well as photography diffi-cult. No additional remains of Sphenopteris philipensis have been recorded since its introduction by Bell (1944).
12 Bell (1944, p. 65) noted the resemblance of Sphenopteris philipensis to Crossotheca kidstonii and Sphenopteris hoeninghausii (= Lyginopteris hoeninghausii). The latter species has often been confused with Crossotheca, and Jongmans (in Crookall 1930, p. 635) regarded Crossotheca kidstonii as synonymous with Crossotheca schatzlarensis, a species that Brousmiche (1982, p. 690) considered synonymous with Crossotheca crepinii. The exhaustive illustration of Crossotheca kidstonii by Jongmans’s (1951) and the revision of Crossotheca crepinii by Brousmiche (1982, 1983) make clear that Crossotheca is morphologically highly variable. This has not always been considered when small fragments were assigned to a species. Bell (1944) did not compare Sphenopteris philipensis with Crossotheca crepinii, but the short last order pinnae with small, simple to trilobate pinnules with a strongly convex lamina fit perfectly with the latter species.
13 I also include in Crossotheca crepinii specimens from the Sydney Basin figured as Sphenopteris aculeata by Bell (1938, pl. XI, fig. 5; pl. XII, fig. 1; 1966, pl. XXV, figs. 2–3). Only one fragmentary specimen (the holotype) was figured and described by Bell (1938). It consists of a rachis of the ante-penultimate order on which a few incomplete pinnae are inserted. Bell’s specimen shows sphenopteroid to pecopteroid, simple as well as lobate pinnules, with three to five rounded lobes, and veins that are deeply immersed within the lamina. Although this specimen is difficult to evaluate from the illustration and I have not been able to reexamine it directly, its general aspect and Bell’s description suggest that it falls within the morphological variation of Crossotheca crepinii. The additional specimens of Sphenopteris aculeata figured by Bell (1966, pl. XXV, figs. 2–3) are better preserved and quite characteristic of Crossotheca crepinii. As with Sphenopteris philipensis, no further records of Sphenopteris aculeata exist.
14 Comparisons. Vegetative pinnules of Crossotheca schatzlarensis show one to three pairs of narrower, more widely extended lobes. The pinnules of Lyginopteris hoeninghausii are also highly variable, from orbicular to obtusely subtriangular, and each has a markedly convex lamina. However, its pinnules are spatulate and truncate, and the broad rachises are always punctate. Eusphenopteris pulchrior, a small-pinnuled pteridosperm, is also superficially similar to Crossotheca crepinii, but is characterized by shorter, more commonly trilobate, slightly larger pinnules.
15 Stratigraphic and geographic distribution. Although widespread, Crossotheca crepinii is an uncommon species. Brousmiche (1982) records it as a characteristic element of lower to middle Westphalian C (Bolsovian) floras. Its total range is from upper Westphalian B (Duckmantian) to the highest Westphalian D (Asturian).
16 Occurrence in the Maritime Provinces, Canada. Cumberland Basin (Nova Scotia): Bell (1944): locality 854 (GSC 5962 ― paratype of Sphenopteris philipensis); locality 860 (GSC 5595 ― fragmentary + GSC 5856 + GSC 5987 ― holotype + GSC 5989; the latter two specimens are figured herein as a single reconstructed fragment ― see Fig. 1d) + GSC 5988 + GSC 5990 ― paratypes of Sphenopteris philipensis); locality 752 (GSC 1659 + GSC 2897 ― both figured as Corynepteris winslovii by Bell 1938) + GSC 2103 + GSC 2111 + GSC 2113 + GSC 2114 ― all figured as Crossotheca boulayi). Bell (1966): locality 752 (GSC 15048 ― figured as Sphenopteris (Crossotheca) boulayi); locality 1314 (GSC 15042 ― figured as Sphenopteris aculeata).
Crossotheca compacta Bell 1938
(Fig. 1c)
17 Remarks. For the sake of completeness, I illustrate part of an immature fertile specimen (Fig. 1c) from the Sydney Basin that is the counterpart of the holotype of Crossotheca compacta Bell (1938, pl. XXXVII, fig. 1). Crossotheca compacta is a species of doubtful utility since immature (closed) fertile remains are difficult to separate at species level. This specimen is identified as nº 1125 in the GSC collections.
Order Urnatopteridales Danzé 1956
Family Urnatopteridaceae Danzé 1956
Genus Zeilleria Kidston 1884
- p 1877 Calymmotheca Stur, p. 255.
- 1883 Calymmatotheca Zeiller, p. 182 (corrected spelling).
- 1884 Zeilleria Kidston, p. 590–591.
- 1924 Zeilleria Kidston, p. 427–429.
- 1956 Zeilleria Kidston; Danzé, p. 73–75, 241–245.
- 1960 Bertrandia Dalinval, p. 186 (acc. to Brousmiche and Laveine 1982).
- 1971 Zeilleria Kidston; Thomas and Crampton, p. 283.
- 1983 Zeilleria Kidston; Brousmiche, p. 197–199.
18 Type.Zeilleria delicatula Kidston 1884.
19 Remarks. The genus Zeilleria was instituted by Kidston (1884) for the fertile parts of three species previously described: Sphenopteris delicatula, Calymmotheca avoldensis and Calymmotheca frenzlii. It is characterized by globular to oval synangia that are situated at the end of lateral veins; these split into four or five sporangia at maturity. Stur’s Calymmotheca refers to cupules, not microsporangiate structures. Kidston (1884) initially believed Zeilleria to be a fern, but later referred it to the pteridosperms as a result of a mistaken interpretation as seed-bearing cupules (Kidston 1911, 1924).
20 Dalinval (1960) introduced the generic name Bertrandia to accommodate Zeilleria avoldensis, the one species with pecopteroid pinnules; all the other species of Zeilleria possess sphenopteroid pinnules. Bertrandia has been treated as a synonym of Zeilleria by subsequent authors (e.g., Brousmiche and Laveine 1982).
Zeilleria avoldensis (Stur 1883) Kidston 1884
(Figs. 2a–e)
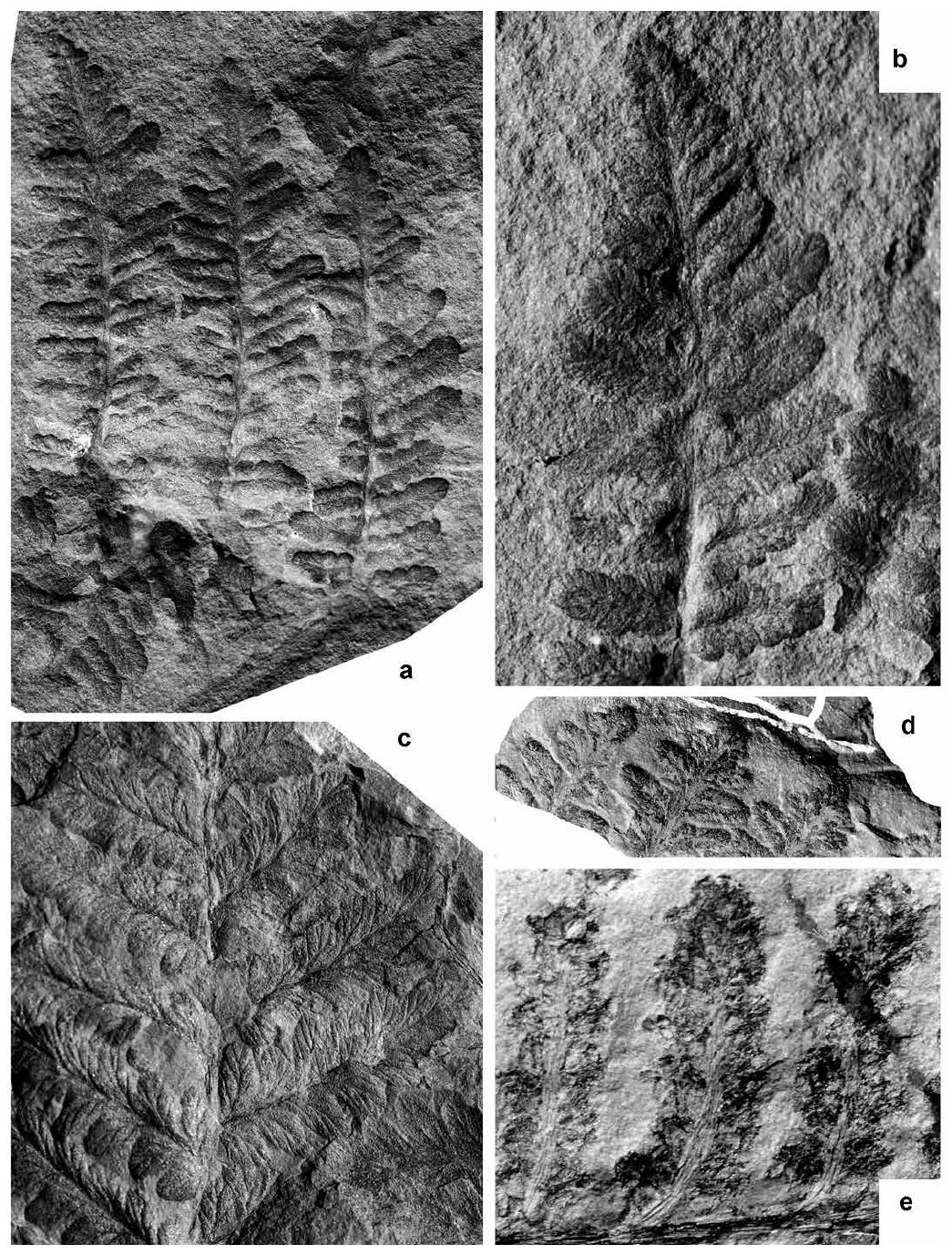 Display large image of Figure 2
Display large image of Figure 2
- * ? 1868 Sphenopteris pilosa Dawson, p. 534, 548, 552, fig. 192F (diagrammatic drawing).
- ? 1871 Callipteris pilosa Dawson, p. 51, pl. XVI, fig. 189–189b (diagrammatic drawings).
- * ? 1871 Pecopteris (Cyathites?) densifolia Dawson, p. 56, pl. XVII, figs. 195–196 (drawings — included by Stopes 1914 in Pecopteris miltonii, and by Bell 1944 in Pecopteris pilosa).
- * 1883 Calymmotheca Avoldensis Stur, p. 171 [803], Figs. 37a–b.
- § 1884 Zeilleria avoldensis (Stur) Kidston, p. 591.
- 1885 Calymmotheca Avoldensis Stur, p. 251–255, Taf. XXXVII, fig. 1; text–figs. 41a–b.
- p 1914 Pecopteris Miltoni (Artis) Brongniart; Stopes, p. 41–44, pl. VII, fig. 14 (photograph of Callipteris pilosa Dawson 1871, pl. XVI, fig. 189), fig. 15, fig. 16 (same as Dawson 1871, pl. XVII, fig. 195); text-figs. 5–6 (drawings); non pl. VII, fig. 17 (same as Dawson 1871, pl. XVII, fig. 196 – indeterminable).
- 1924 Zeilleria Avoldensis (Stur) Kidston; Kidston, p. 432–436, pl. XCVIII, figs. 1–2, figs. 3–6 (spores), fig. 8; pl. XCIX, figs. 1–2b, fig. 8.
- 1927 Zeilleria avoldensis (Stur) Kidston; Corsin, p. 28–35, pl. I, figs. 1–7; pl. II, figs. 2, 5–6.
- 1928 Zeilleria avoldensis (Stur) Kidston; Jongmans, p. 21, pl. 14, figs. 3–4.
- ? 1938 Zeilleria avoldensis (Stur) Kidston; Bell, p. 35, pl. XXII, fig. 5 (difficult to judge from the illustration).
- 1938 Pecopteris clarkii Lesquereux; Bell, p. 82, pl. LXXXIII, fig. 3; pl. LXXXIV, fig. 1.
- v p 1944 Pecopteris pilosa (Dawson) Bell, p. 85–86, pl. XIII, figs. 1, 5; pl. XXXIX, figs. 1–4; pl. XL, fig. 2, figs. 3–4 (herein Fig. 2d), fig. 5; pl. XLI, figs. 1, 4; pl. XLII, figs. 1, 6 (partially refigured herein as Fig. 2e), fig. 3; pl. XLIII; pl. XLIV (? – specimen from New Brunswick not available for revision that raises some doubts about its attribution to Zeilleria avoldensis).
- * p 1951 Pecopteris tenuinervosa Corsin, p. 310–311, pl. CLXIV, fig. 3; non pl. CLXIV, fig. 4 (= Pecopteris micromiltonii) (acc. to Brousmiche and Laveine 1982).
- 1951 Pecopteris cf. avoldensis (Stur) Corsin; Corsin, p. 355–356, pl. CLXXXIV, figs. 1–2a.
- * 1958 Zeilleria lignyi Stockmans and Willière, pl. VII, figs. 1–4 (well illustrated but only diagnosed briefly in the plate explanation); pl. VIII, figs. 1–4 (fertile specimens assigned to Bertrandia by Dalinval 1960) (acc. to Álvarez-Vázquez 1995).
- 1960 Pecopteris (Bertrandia) avoldensis (Stur) Dalinval (including synonymy), p. 185–194, pl. LVII, figs. 1–3a; pl. LVIII, figs. 1–3a; pl. LIX, figs. 1–3a; pl. LX, figs. 1–4a; pl. LXI, figs. 1–3a.
- p 1960 Pecopteris (Asterotheca) miltoni (Artis) Zeiller; Dalinval, pl. 33, figs. 1–3a; pl. 34, figs. 1–3a; pl. 35, figs. 1–2a; pl. 36, figs. 1–4a; pl. 40, fig. 1 (with doubt); non p. 133–149, non pl. 32, figs. 1–1b (= Pecopteris pseudovestita); non pl. 37, figs. 1–1a (= Pecopteris cf. miltonii); non pl. 37, figs. 2–2a (= Pecopteris sp.); non pl. 37, fig. 3 (= Pecopteris miltonii); pl. 38, figs. 1–2a (= Peco pteris miltonii); pl. 39, figs. 1–2b (= Pecopteris cf. miltonii); non pl. 40, figs. 2–3 (fertile pinnules of Asterotheca-type different to that shown by Pecopteris miltonii) (acc. to Wagner and Álvarez-Vázquez 2016).
- 1960 Pecopteris; Gillespie and Latimer, p. 53, pl. 17, fig. 4.
- v 1965 Pecopteris cf. miltoni (Artis) Brongniart; Stock-mans and Willière, pl. IV, figs. 2–3.
- 1966 Zeilleria avoldensis (Stur) Kidston; Bell, p. 28, pl. XIII, fig. 4.
- * 1966 Asterotheca praecursor Stockmans and Willière, p. 8–9, pl. III, figs. 1–4a (compared with Bertrandia avoldensis and Pecopteris miltonii by Stockmans and Willière).
- * 1966 Asterotheca inferior Stockmans and Willière, p. 9, pl. IV, figs. 1–3a (fertile specimens from the same locality as those of Asterotheca praecursor).
- 1966 Pecopteris; Gillespie et al., p. 100, pl. 30, fig. 2 (same as Gillespie and Latimer 1960, pl. 17, fig. 4).
- 1970 Pecopteris avoldensis (Stur) Corsin; Laveine, p. 282–286, pl. 37, figs. 1–1a (same as Dalinval 1960, pl. LX, fig. 3); figs. 1b–k (spores); pl. 38, figs. 1–2a, 3–3a (with cf.); figs. 2b–f, 3b–g (spores).
- * 1971 Pecopteris alloiophylla Doubinger and Germer, p. 74, Abb. 2, Taf. 21, figs. 1a–b (acc. to Brousmiche and Laveine 1982).
- * 1971 Pecopteris engyphylla Doubinger and Germer, p. 75–76, Abb. 5, Taf. 21, figs. 4a–b (acc. to Brousmiche and Laveine 1982).
- * 1971 Pecopteris pachyneura Doubinger and Germer, p. 81, Abb. 12, Taf. 23, figs. 3a–b (acc. to Brousmiche and Laveine 1982).
- * 1971 Pecopteris pycnoneura Doubinger and Germer, p. 83, Abb. 14, Taf. 24, figs. 1a–c (acc. to Brousmiche and Laveine 1982).
- * 1971 Pecopteris brachyphylla Doubinger and Germer, p. 83, Abb. 15, Taf. 24, figs. 2a–b (acc. to Brousmiche and Laveine 1982).
- 1971 Zeilleria avoldensis (Stur) Kidston; Thomas and Crampton, p. 283–294, pl. I, figs. A–G; pl. II, figs. A–I (spores); text-figs. 1–3 (drawings).
- T 1982 Pecopteris (Zeilleria) avoldensis (Stur) Kidston; Brousmiche and Laveine, p. 52–58 (including synonymy), pl. I, figs. 1–1b (neotype as designated by Brousmiche 1983), figs. 2–3b; pl. II, figs. 1–8a, figs. 9–9a (same as Corsin 1951, pl. CLXIV, fig. 3); pl. III, figs. 1–8a; pl. IV, figs. 1–1a, 11–11a, figs. 2–10, 12 (spores); text-figs. 1A–2.
- 1983 Zeilleria avoldensis (Stur) Kidston; Brousmiche, p. 215–217, pl. 52, figs. 1–1a (same as Corsin 1951, pl. CLXIV, fig. 3), figs. 2–8a.
- 1985 Sphenopteris sp. A; Lyons et al., p. 228, 234, pl. XI, fig. e.
- v 2001b Zeilleria pilosa (Dawson) comb. nov. Wagner, p. 95, fig. 46 (same as figured as Zeilleria avoldensis by Álvarez-Vázquez 1995, lám. 54, fig. 1).
- 2002 Zeilleria cf. avoldensis (Stur) Kidston; Blake et al., p. 268, 291, 317, pl. XVII, fig. 5 (fertile).
- v 2010 Zeilleria avoldensis (Stur) Kidston; Wagner and Álvarez-Vázquez, p. 257, 266, 268, 270, 307.
21 Description. Frond at least tripinnate. Antepenultimate rachis straight, longitudinally striate, ca. 2 mm wide. Pinnae of penultimate order alternate, probably lanceolate (incompletely preserved), touching laterally or slightly overlapping. Dimensions: at least 70 mm long and 25 mm broad. Penultimate rachis inserted at narrow angle, rounded, straight, longitudinally striate, ca. 0.4–0.5 mm wide. Last order pin-nae alternate, lanceolate, generally touching laterally, with a rounded, poorly individualized terminal; dimensions: 10–30 mm long and 5–10 mm broad; length/breadth ratio = 2–3. Last order rachis rounded, straight, longitudinally striate, ca. 0.1 mm wide. Pinnules alternate, ovate to subrectangular, inserted slightly obliquely and broadly attached (pecopteroid). Fully developed pinnules subrectangular, with a slightly constricted base, lobed margins and rounded apex. Dimensions: 4–6 mm long and 1.5–2 mm broad; length/breadth ratio = 2.6–3. Smaller, less individualized pinnules ovate, united for up to one half of their length. Dimensions: 3–4 mm long and 1.5–2 mm broad; length/ breadth ratio = 2. Nervation well-marked, deeply embedded in the convex lamina. Midrib slightly decurrent and extending into the pinnule apex; lateral veins once or twice forked. Fertile pinnules similar in shape to the vegetative ones. Immature synangia oval, placed at the end of lateral veins. Mature synangia extend beyond a slightly reduced lamina; at full maturity these split into four or five sporangia.
22 Remarks. Stopes (1914) figured photographically the specimens of Sphenopteris pilosa that were illustrated by diagrammatic drawings by Dawson (1868, 1871). She included this species in synonymy with Pecopteris miltonii, apparently in error (Wagner and Álvarez-Vázquez 2016). Stopes (1914) also synonymized Dawson’s (1871) Pecopteris densifolia with Pecopteris miltonii. Bell (1944, p. 85) agreed with Stopes (1914) that the vegetative remains of Sphenopteris pilosa resemble those of Pecopteris miltonii, but he regarded the synangia as different. He thus rejected Stopes’s synonymy and reinstated Dawson’s species as Pecopteris pilosa. The correct identification of Pecopteris miltonii is a problem that was dealt with by Wagner and Álvarez-Vázquez (2016) based on a redescription of material from the type area of the species in South Yorkshire, England, and other European localities. Wagner and Álvarez-Vázquez (2016) indicated in their synonymy list that specimens belonging to Zeilleria avoldensis have often been mistaken for Pecopteris miltonii.
23 The type material of Sphenopteris pilosa, figured and described by Dawson (1868, 1871) and Stopes (1914), originated from the Fern Ledges locality at Saint John, New Brunswick. Additional specimens from Nova Scotia, including fertile remains, were figured and described as Pecopteris pilosa by Bell (1944). Bell’s specimens have been re-examined in present study and compared with European material of Zeilleria avoldensis. At least one of the fertile specimens from Nova Scotia figured by Bell (1944, pl. XXXIX, fig. 3; pl. XL, fig. 2) shows fully opened synangia situated at the tip of pinnule lobes. This specimen closely resembles that figured by Thomas and Crampton (1971) from lower Bolsovian strata of Great Britain. Bell’s specimens should be included in Zeilleria avoldensis.
24 Zeilleria avoldensis was revised by Brousmiche and Laveine (1982), who clarified many misidentifications and included several species in synonymy. European specialists consistently ignored the Canadian material until Wagner (2001b) introduced the new combination Zeilleria pilosa in a list of species of the Peñarroya-Belmez-Espiel coalfield, southwestern Spain, thus placing Zeilleria avoldensis in synonymy with Sphenopteris pilosa.
25 I have not been able to review Dawson’s types of Sphenopteris pilosa and Pecopteris densifolia; thus here they have been included only questionably in synonymy with Zeilleria avoldensis. If future work confirms the synonymy of Zeilleria avoldensis with one or more of the earlier names, a formal proposal should be made to conserve Zeilleria avoldensis. The very small fragments figured by Dawson (1868) as Sphenopteris pilosa contrast with the large, well-preserved fertile specimen figured and described by Stur (1885, Taf. XXXVII, fig. 1) on which Zeilleria avoldensis is based. Most important is the widespread use of Zeilleria avoldensis in the paleobotanical literature, whereas Zeilleria pilosa has been rarely used.
26 Comparisons.Zeilleria avoldensis is easily distinguished from the other species of Zeilleria by its thick, convex lamina, and the presence of pecopteroid pinnules.
27 Stratigraphic and geographic distribution.Zeilleria avoldensis is known from numerous localities in Europe, where it has been commonly recorded, although not always under this name (see synonymy list). It is long-ranging, from (Yeadonian?) Langsettian to middle Asturian. Stur’s type material originated from middle Bolsovian strata of Saar-Lorraine. Corsin’s (1951) Pecopteris tenuinervosa and the five species distinguished by Doubinger and Germer (1971) all came from different levels of Bolsovian and Asturian strata in Saar-Lorraine. The type materials of Zeilleria lignyi, Asterotheca inferior and Asterotheca praecursor came from upper Langsettian strata in Belgium. Wagner and Álvarez-Vázquez (2010) recorded the species from various levels of Langsettian to lower Asturian strata in different parts of the Iberian Peninsula. The few records of Zeilleria avoldensis from the USA may not properly reflect its occurrence in North America as it seems to be one of the more common fern species in the Maritime Provinces of Canada.
28 Occurrence in the Maritime Provinces, Canada. Cumberland Basin (Nova Scotia): Bell (1944): locality 999 (GSC 8205 + GSC 8596 + GSC 8597 ― fragmentary + five pieces without catalogue number ― together with Alethopteris urophylla, Annularia ramosa and Asterophyllites sp.); locality 1031 (GSC 8208 ― fertile + one piece with damaged catalogue number); locality 1086 (GSC 9986 ― together with Calamites carinatus); locality 1088 (GSC 10116 ― with Calamites sp. and Asterophyllites sp.); locality 1010 (GSC 8661); locality 1342 (GSC 8218 + GSC 10970); locality 1389 (GSC 8211 + GSC 10010 + GSC 10011); locality 1406 (fertile specimens ― two pieces, part and counterpart, without catalogue number); locality 2253 (two pieces without catalogue number ― fragmentary; together with Laveineopteris polymorpha); locality 3115 (one piece without catalogue number ― fragmentary); locality 3995 (one piece without a catalogue number ― together with Samaropsis sp.); locality 5868 (three pieces without catalogue number ― fragmentary and poorly preserved). Collected in 1960: locality 5644 (one piece without catalogue number). Saint John (New Brunswick): Bell (1944): locality 203 (one piece without catalogue number ― fragmentary); locality 351 (one piece ― fragmentary); locality 352 (one piece without catalogue number); locality 559 (GSC 8201 ― fragmentary); locality 785 (GSC 8204). New Brunswick Museum collection: NBMG 1842 (fragmentary; with cf.) + NBMG 10532 (labelled as Sphenopteris pilosa) + NBMG 12049/1 (together with Dorycordaites palmaeformis) + NBMG 12049/2 + NBMG 12049/3 + NBMG 12049/4 (together with Dorycordaites palmaeformis). Sydney Basin (Nova Scotia): Bell (1938): locality 480 (GSC 1669); locality 518 (GSC 3057 + GSC 3058). Bell (1966): locality 480 (GSC 15049).
29 Occurrence in the United States. Alabama: Lyons et al. (1985). West Virginia: Gillespie and Latimer (1960), Gillespie et al. (1966), Blake et al. (2002).
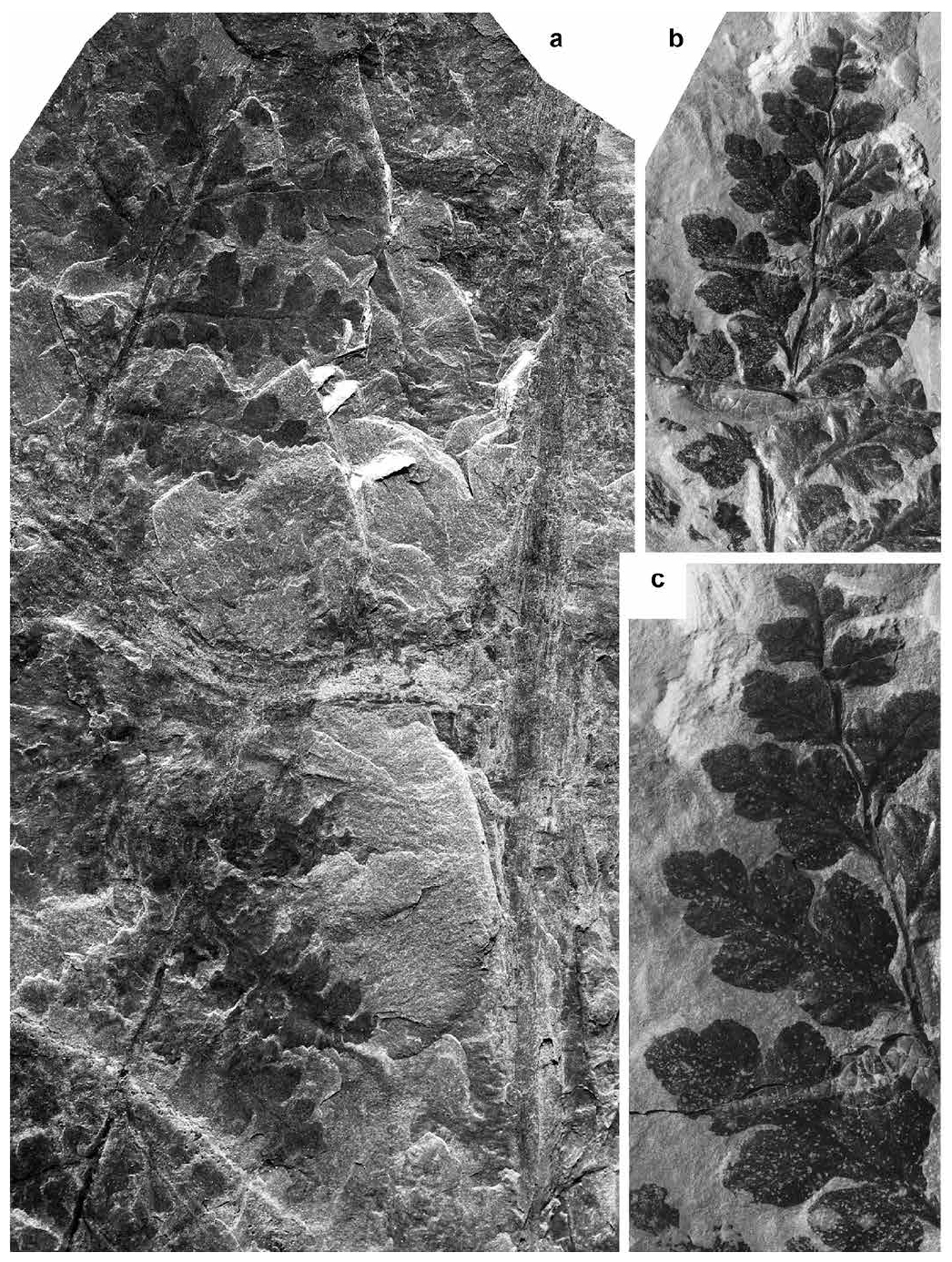
Zeilleria frenzlii (Stur 1883) Kidston 1884
(Figs. 3a–d)
 Display large image of Figure 3
Display large image of Figure 3
- * 1883 Calymmotheca Frenzli Stur, p. 172 [804], Figs. 38a–b.
- § 1884 Zeilleria Frenzlii (Stur) Kidston, p. 591.
- 1885 Calymmotheca Frenzli Stur, p. 268–272, Taf. XXXVII, figs. 2–3; Taf. XXXVIII, fig. 3; text-figs. 42a–b.
- * 1885 Calymmotheca Walteri Stur, p. 263–264, Taf. XXXVI, fig. 4 (specimen from the same horizon as two of the three other specimens figured by Stur 1885 as Calymmotheca frenzlii).
- 1914 Sphenopteris (Zeilleria) Frenzli (Stur) Kidston; Carpentier and Depape, p. 307–310, pl. IV, figs. 1–4 (magnificent fertile specimens), fig. 5 (sporangia); pl. VI, fig. 1; text-figs. 1–2.
- * 1937 Hymenotheca globulifera Němejc, p. 9, pl. II, figs. 5, 6 (Němejc made comparisons with Zeilleria frenzlii and Crossotheca schatzlarensis).
- 1938 Zeilleria frenzli (Stur) Kidston; Bell, p. 34, pl. XX, figs. 2–3; pl. XXI, fig. 2.
- p 1938 Hymenotheca dathei Potonié; Bell, p. 36, pl. XX, fig. 4; pl. XXI, fig. 1; non pl. XXIII, figs. 4–5 (difficult to judge, but the specimen shows pinnules with a broader lamina than Zeilleria frenzlii).
- 1941 Sphenopteris cf. sancti felicis (Stur) Rydzewski; Gothan, p. 11–12, Taf. 49, fig. 3 (Stur’s Diplotmema sanctifelicis is a pteridosperm, not a fern).
- T 1943 Zeilleria frenzli (Stur) Kidston; Guthörl, p. 144, Taf. XXIX, Abb. 2A–2B, Abb. 3A–3B (neotype designated by Brousmiche 1983).
- v 1944 Sphenopteris dixoni Kidston; Bell, p. 67, pl. XIV, figs. 2–3 (fragmentary).
- * v 1944 Sphenopteris fletcheri Bell, p. 68, pl. XVI, fig. 2 (paratype); pl. XVI, fig. 6 (holotype — Figs. 3a–b herein).
- v 1944 Zeilleria frenzli (Stur) Kidston, Bell, p. 73, pl. XXII, figs. 1–2; pl. LXXVII, fig. 6; pl. LXXVIII (partially refigured in Figs. 3c–d); pl. LXXIX, figs. 2–3.
- 1958 Zeilleria frenzli (Stur) Kidston; Langford, p. 282, Fig. 518; Fig. 519a (drawing).
- v 1965 Zeilleria frenzli (Stur) Kidston; Wagner, p. 88–90, pl. 11, figs. 17–18.
- 1969 Zeilleria frenzli (Stur) Kidston; Skog et al., Figs. 10–11; Fig.12 (spores).
- 1983 Zeilleria frenzli (Stur) Kidston; Brousmiche, p. 205–212 (including synonymy), pl. 49, figs. 1–5, figs. 6–6a (same as Álvarez Ramis et al. 1978, Taf. 9, fig. 1); pl. 50, figs. 1–4; text-figs. 61A1–E3.
- p 1985 Zeilleria delicatula Kidston; Gillespie and Craw-ford, p. 252, pl. II, fig. 4 (?), fig. 5 (fertile); non pl. II, fig. 3 (= Renaultia schatzlarensis).
- p 2002 Zeilleria frenzlii (Stur) Kidston; Blake et al., p. 268, 291, pl. XVII, fig. 2; non pl. XVII, fig. 3 (= Renaultia schatzlarensis).
- v 2010 Zeilleria frenzlii (Stur) Kidston; Wagner and Álvarez-Vázquez, p. 257, 266.
Excludenda:
- 2017 cf. Zeilleria frenzlii (Stur) Kidston; DiMichele et al., p. 53–54, 55, figs. 31.10–31.11 (small, fragmentary specimen resembling Germera mendescorreae).
30 Description. Frond at least tripinnate. Penultimate rachis straight, flat, longitudinally striate, ca. 1 mm wide. Last order pinnae lanceolate, alternate or subopposite, laterally touching or slightly overlapping. Dimensions: 10–40 mm long and 6–13 mm broad; length/breadth ratio = 1.6–3. Last order rachis inserted at 45–80°, straight, flat, longitudinally striate, narrowly winged due to decurrent, confluent pinnule bases, ca. 0.7 mm wide. Pinnules alternate or sub-opposite, obliquely inserted; less developed pinnules (in the apical parts of pinnae) consist of one or two linear segments with slightly pointed apices (which may appear subrounded as a result of imperfect preservation). More fully developed pinnules are deeply incised and split into five to six parallel-sided, narrow lobes, with acute apices. Dimensions: 5–12 mm long and 1.5–5 mm broad; length/breadth ratio = 2.4–3.25. A single, relatively thick vein enters each lobe extending into the apex. Fertile pinnules with narrower lobes than those of vegetative ones. Sporangia globular, relatively large (1–1.2 mm), placed at the end of lobes; when fully mature, the sporangia split into four segments.
31 Remarks. The thin, almost filiform segments into which the pinnules of Zeilleria frenzlii are split clearly distinguish this species, especially when sporangia are present. Fertile specimens were well illustrated by Bell (1944, pl. XXII, fig. 1; pl. LXXVII, fig. 6; pl. LXXVIII; pl. LXXIX, figs. 2–3).
32 Bell (1944, p. 68, pl. XVI, figs. 2, 6) introduced a new species, Sphenopteris fletcheri, from two localities in the Cumberland Basin of Nova Scotia. He compared it with two pteridosperm taxa, Sphenopteris alata (= Palmatopteris alata) and Diplotmema furcatum (= Palmatopteris furcata), although the comparison now appears to be irrelevant. Bell apparently overlooked Zeilleria frenzlii, which I regard as the senior synonym of Sphenopteris fletcheri. No subsequent records of Sphenopteris fletcheri are known.
33 I also include in Zeilleria frenzlii the fragmentary specimen figured by Bell (1944, pl. XIV, figs. 2–3) as Sphenopteris dixonii. This specimen shows slightly decurrent, linear, trilobed pinnules with narrow pointed lobes, which are essentially identical to those of Stur’s species. Kidston’s (1917) Sphenopteris dixonii possesses smaller, Corynepteris-type pinnules inserted in parallel-sided pinnae; these pinnules are cuneate and show two, or rarely three, blunt segments on the upper margin.
34 Comparisons. Vegetative pinnules of Zeilleria schaumburglippeana have broader lamina and obtusely pointed lobes. According to Brousmiche (1983), Zeilleria frenzlii has pinnules that are one and a half times longer than those of Zeilleria schaumburglippeana at the same stage of differentiation. Vegetative pinnules of Germera brousmicheae sp. nov. (see below) also possess up to three pairs of parallel-sided lobes, but these display a much wider lamina and markedly more rounded or tongue-shaped apices.
35 Stratigraphic and geographic distribution.Zeilleria frenzlii is a widespread species, ranging from Langsettian to upper Bolsovian. The holotype is from lower Bolsovian strata of Saar-Lorraine, from where Brousmiche (1983) noted that the species is common throughout the Bolsovian. Němejc’s (1937) Hymenotheca globulifera originated from Bolsovian strata of the Lubná group of coals, Upper Radnice, Kladno Basin, Bohemia, Czech Republic.
36 Occurrence in the Maritime Provinces, Canada. Cumberland Basin (Nova Scotia): Bell (1944): locality 1039 (GSC 9064 ― figured as Sphenopteris fletcheri + GSC 9224 ― holotype of Sphenopteris fletcheri; together with Cyperites sp.); locality 1086 (GSC 10038 + GSC 10052 + GSC 10054 + GSC 10056 ― all fertile remains); locality 1141 = 666 (GSC 10243 + GSC 10244 ― part and counterpart; figured as Sphenopteris dixonii by Bell); locality 1362 (GSC 10976 + one fragmentary specimen without catalogue number); locality 1462 (one piece without catalogue number ― together with rootlets); locality 1517 (one piece from a borehole without catalogue number); locality 2253 (one piece without catalogue number ― together with Laveineopteris polymorpha, Renaultia sp. and Dorycordaites palmaeformis). Sydney Basin (Nova Scotia): Bell (1938): locality 482 (GSC 1641); locality 484 (GSC 1639).
37 Occurrence in the United States. Georgia: Gillespie and Crawford (1985); Blake et al. (2002). Illinois: Langford (1958).
Zeilleria hymenophylloides Kidston 1924
(Figs. 4a–b; Figs. 5a–b)
 Display large image of Figure 4
Display large image of Figure 4
 Display large image of Figure 5
Display large image of Figure 5
- p 1916 Sphenopteris Kayi Arber, pl. 2, fig. 3; non p. 142–143, pl. 2, fig. 6 (excluding the holotype).
- * 1924 Zeilleria hymenophylloides Kidston, p. 439–440, pl. XCVII, fig. 1 (lectotype designated by Brousmiche 1983), pl. XCVII, figs. 2–2a; pl. XCIX, figs. 9–11.
- p 1928 Zeilleria hymenophylloides Kidston; Němejc, p. 1–2, pl. I, fig. 1, figs. 2–4 (sporangia); non pl. II, fig. 1 (difficult to judge from the illustration).
- 1933 Zeilleria fructification; Arnold, p. 822, figs. 1–2 (drawings), fig. 3 (spores).
- * 1937 Zeilleria stellata Arnold, p. 4, pl. 1, fig. 4 (same as Arnold 1933).
- 1941 Sphenopteris (Zeilleria) hymenophylloides (Kidston) Gothan, p. 17, Taf. 52, figs. 4–5a; Taf. 53, fig. 2.
- v 1944 Sphenopterisb(Zeilleria) hymenophylloides (Kidston) Gothan; Bell, p. 72–73, pl. XIII, fig. 6; pl. XX, fig. 3 (herein Fig. 4b); pl. XXII, fig. 3 (poorly preserved).
- 1944 Sphenopteris moyseyi Kidston; Bell, p. 66, pl. XIV, fig. 4.
- v 1944 Zeilleria schaumberglippeana (sic) (Stur) Zeiller; Bell, p. 73–74, pl. XXIII, figs. 4–5; pl. XXIV, fig. 3.
- ? 1958 Zeilleria delicatula Kidston; Langford, p. 282, Fig. 519-2a (drawing); Fig. 520 (poor figure).
- 1983 Zeilleria hymenophylloides Kidston; Brousmiche, p. 213–215 (including synonymy), pl. 51, figs. 1–3a (same as Kidston 1924, pl. XCIX, figs. 9–11), figs. 4–5, figs. 6–8 (as Zeilleria cf. hymenophylloides).
- v 1987 Zeilleria hymenophylloides Kidston; Wagner, lám. 8, figs. 1–2.
- 1988 Zeilleria hymenophylloides Kidston; Amerom, p. 97–102, pl 1, figs. 1–7; pl. 2, figs. 1–6; text-figs. 1–4.
- v 1995 Zeilleria hymenophylloides Kidston; Álvarez- Vázquez, p. 179–181 (including synonymy), lám. 54, fig. 1 (together with Zeilleria avoldensis and Paripteris sp.); lám. 56, fig. 3; lám. 58, fig. 1 (same as Wagner 1987, lám. 8, fig. 1, at natural size), fig. 3; lám. 59, fig. 1 (same as Wagner 1987, lám. 8, fig. 1, enlarged), figs. 2–3; pl. 60, figs. 1–5.
- v 2001b Zeilleria hymenophylloides Kidston; Wagner, p. 95, fig. 44 (same as Wagner 1987, lám. 8, fig. 1, and Álvarez-Vázquez, 1995, lám. 58, fig. 1; lám. 59, fig. 1), fig. 45.
- v 2010 Zeilleria hymenophylloides Kidston; Wagner and Álvarez-Vázquez, p. 254, 255, 257, 266, 307, pl. V, fig. 1.
38 Description. Frond at least tripinnate. Antepenultimate rachis straight, smooth, ca. 2 mm wide. Pinnae of penultimate order alternate, well spaced, apparently subrectangular (always incomplete), at least 45 mm long and 20 mm broad. Penultimate rachis inserted at 45–50°, straight, smooth, ca. 0.7 mm wide. Last order pinnae alternate, closely spaced but not touching, subrectangular, elongate, 10–20 mm long and 3–6 mm broad; length/breadth ratio ≈ 3.3. Last order rachis inserted at 75–80°, straight, smooth or showing a faint longitudinal striation, ca. 0.2–0.3 mm wide. Pinnules alternate, decurrent, attached to the rachis by a wide footstalk; general oultine ovate and divided into two to five simple or bifid pairs of lobes with obtusely pointed apices; terminal lobe simple of bifid. Dimensions: 3–5 mm long and 2–3 mm broad; length/breadth ratio = 1.5–1.6. Lamina thick, convex. Midrib thin, slightly flexuous, once or twice bifurcate, remaining well developed to near the pinnule apex.
39 Remarks. Although fragmentary and poorly preserved, I agree that the two specimens illustrated by Bell (1944, pl. XIII, fig. 6; pl. XX, fig. 3; pl. XXII, fig. 3) should be assigned to Sphenopteris (Zeilleria) hymenophylloides. The better preserved specimen figured as Sphenopteris moyseyi by Bell (1944, pl. XIV, fig. 4), originating from the same locality as one of Bell’s specimens identified as Zeilleria hymenophylloides (locality 2567), may also belong to this species.
40 The two specimens figured by Bell (1944, pl. XXIII, figs. 4–5; pl. XXIV, fig. 3) as Zeilleria schaumburglippeana were incorporated by Brousmiche (1983, p. 353) in her synonymy list for Zeilleria hymenophylloides. However, she emphasized the poor quality of Bell’s illustrations and preferred to attribute the material to Sphenopteris incertae sedis. Although the delicate lamina is poorly preserved, Bell’s specimens clearly possess fertile, decurrent, ovate pinnules with simple or bifid, obtusely pointed lobes. Re-examination of Bell’s specimens, along with thirteen additional from Bell’s locality 788, confirms that these specimens should be attributed to Zeilleria hymenophylloides.
41 Zeilleria hymenophylloides has been figured and described in detail by Brousmiche (1983), Amerom (1988) and Álvarez-Vázquez (1995) from various parts of western Europe.
42 Comparisons. The pecopteroid aspect and thick, sub-rectangular lamina of vegetative pinnules of Zeilleria avoldensis make this species easily distinguishable from Zeilleria hymenophylloides. The latter species shows more sphenopteroid, ovate pinnules divided into simple or bifid lobes; the pinnules are attached to the rachis by a wide foot-stalk. Zeilleria frenzlii differs in having longer, more narrowly winged pinnules, divided into more filiform lobes.
43 Stratigraphic and geographic distribution.Zeilleria hymenophylloides is a rarely recorded species. Álvarez-Vázquez (1995) gives its total range as middle Langsettian to middle Bolsovian. Kidston’s holotype originated from upper Langsettian strata of the Lancashire coalfield of England. Brousmiche (1983) recorded the species (as Zeilleria cf. hymenophylloides) from middle Westphalian C (Bolsovian) of Saar-Lorraine. Amerom’s (1988) specimens are from Langsettian strata of South Limburg, the Netherlands.
44 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin: Bell (1944): locality 1031 (one poorly preserved specimen without catalogue number ― together with Cordaites sp.); locality 3109 (GSC 5596 + five pieces without catalogue number). Saint John (New Brunswick): Bell (1944): locality 701 (one poorly preserved specimen without catalogue number); locality 788 (GSC 10978 ― fertile specimen figured as Zeilleria schaumburglippeana by Bell + fourteen pieces without catalogue number); locality 808 (one piece without catalogue number ― fragmentary and poorly preserved); locality 1498 (two pieces without catalogue number); locality 2567 (GSC 9217 + GSC 9375 ― figured as Sphenopteris moyseyi + two poorly preserved specimens without catalogue number); locality 2569 (GSC 5598 + two pieces without catalogue number ― fragmentary and poorly preserved). Fundy Geological Museum: FGM 997 GF 31.14 (fragmentary and poorly preserved ― together with Laveineopteris polymorpha and Dorycordaites palmaeformis).
45 Occurrence in the United States. Illinois: Langford (1958). Michigan: Arnold (1933, 1937).
Genus Renaultia Zeiller 1883
- 1883 Renaultia Zeiller, p. 185 (August).
- 1883 Hapalopteris Stur, p. 660 [28] (December).
- 1956 Renaultia Zeiller; Danzé, p. 69–70, 171–175.
- 1983 Renaultia Zeiller; Brousmiche, p. 138–140.
46 Type.Renaultia chaerophylloides (Brongniart 1836) Zeiller 1883.
47 Remarks.Renaultia encompasses small-pinnuled ferns characterized by a delicate, more or less reduced lamina, and sporangia situated at the vein endings and either single or arranged in groups of two to five. Zeiller (1883, August) introduced Renaultia to incorporate fertile remains previously assigned to Pecopteris chaerophylloides. Almost simultaneously, Stur (1883, December) proposed Hapalopteris for Hapalopteris typica, a synonym of Renaultia chaerophylloides.
Renaultia footneri (Marrat 1872) Kidston 1893
(Fig. 6)
 Display large image of Figure 6
Display large image of Figure 6
- * 1872 Sphenopteris Footneri Marrat, p. 94, pl. VIII, figs. 2–3.
- T 1889 Sphenopteris Footneri Marrat; Kidston, p. 406–407, pl. II, figs. 3–3b.
- § 1893 Renaultia footneri (Marrat) Kidston, p. 104.
- T 1924 Sphenopteris Footneri Marrat; Kidston, p. 117–120, pl. XXIV, figs. 1–1b (lectotype designated by Brousmiche 1983), figs. 2–6; pl. XXV, fig. 5.
- v 1944 Sphenopteris (Renaultia) gracilis (Brongniart) Zeiller; Bell, p. 72, pl. XX, figs. 1–2 (herein Fig. 6).
- 1956 Sphenopteris footneri Marrat; Danzé, p. 444–448, pl. LXXII, figs. 1–3; pl. LXXIV, figs. 1–1b.
- T 1981 Sphenopteris footneri Marrat; Cleal, p. 15, pl. 3, fig. 2 (wrongly cited as fig. 1 in the plate caption); pl. 5, fig. 3 (lectotype), fig. 4 (syntype).
- 1983 Renaultia footneri (Marrat) Kidston; Brousmiche, p. 154–158, pl. 37, figs. 1–4a.
- T 1992 Renaultia footneri (Marrat) Kidston; Cleal, p. 534, Fig. 9 (lectotype).
- v 2010 Renaultia footneri (Marrat) Kidston; Wagner and Álvarez-Vázquez, p. 257, 266.
48 Description. Frond at least tripinnate. Pinnae of antepenultimate order alternate, subtriangular, always incomplete. Antepenultimate rachis inserted at 80–90°, straight, rounded, longitudinally striate, up to 1 mm wide. Pinnae of penultimate order alternate, subtriangular, elongate, always incomplete. Penultimate rachis inserted at 75–80°, slightly flexuous, rounded, smooth, up to 0.4 mm wide. Last order pinnae alternate, distant; subtriangular, elongate, with apical angle of 45–50°. Dimensions: 25–30 mm long and 15 mm wide (at the base); length/breadth ratio = 1.6–2. Last order rachis inserted at 80–90°, slightly flexuous, rounded, smooth, ca. 0.2 mm wide. Pinnules alternate, decurrent, with a thin, flat lamina. Less developed pinnules inserted at 45–50° by the entire base, confluent; subtriangular, equilateral, simple or slightly lobed, with a contracted anadromous margin. Well-developed pinnules inserted at 80–90°, sub-triangular, composed by three or four pairs of lobes, separated by shallow sinuses and with two to four pointed teeth in each lobe. Dimensions: 4–8 mm long and 2–4 mm wide; length/breadth ratio = 2. Veins deeply embedded in the lamina. Midrib thin, slightly flexuous and strongly marked, remaining well developed to near the apex. The midrib gives off a thin lateral vein to each lobe, once or twice forked at wide angles. Fertile pinnules similar to the vegetative ones. Immature synangia small, oval, placed at the end of lateral veins.
49 Remarks. Bell (1944, pl. XX, figs. 1–2) figured as Sphenopteris (Renaultia) gracilis a single specimen from Springhill, Nova Scotia. His illustrations are difficult to evaluate, and later authors consistently ignored Bell’s record (e.g., Brousmiche 1983, who discussed Renaultia gracilis in detail but did not refer to the Canadian material). Re-examination of Bell’s specimen and comparison with the holotype of Renaultia gracilis (Brongniart 1830, pl. 54, figs. 2a–b), as figured photographically by Brousmiche (1983, pl. 38, figs. 1–1b), suggest that Bell’s identification was incorrect. Well-developed pinnules of Renaultia gracilis are subrectangular and composed of three pairs of subangular lobes that are separated by relatively deep sinuses. Bell’s (1944) specimen has subtriangular pinnules with three to four pairs of lobes with two to four pointed teeth in each lobe and separated by shallow sinuses. A comparison with type material of Renaultia footneri (Kidston 1889, 1924; Cleal 1981, 1992), suggests that Bell’s material belongs to the latter species.
50 Comparisons.Renaultia gracilis has subrectangular, elongate pinnules, with up to three pairs of more angular lobes. At the same stage of differentiation, pinnules of Renaultia footneri are shorter, with a lower length/breadth ratio. Well-developed pinnules of Renaultia rotundifolia are also subtriangular and composed of three or four pairs of lobes separated by shallow sinuses. However, Renaultia rotundifolia has pinnules that are less elongate, larger, and have a smaller length/breadth ratio. At the same stage of differentiation, pinnules ofRenaultia chaerophylloides are larger and possess more acute lobes. Both Renaultia footneri and Renaultia chaerophylloides show the first catadromous pinnule smaller than the anadromous one, but in Renaultia chaerophylloides it is markedly smaller and less differentiated. Sphenopteris rutaefolia also possesses subtriangular pinnules, but these are more equidimensional, with a smaller length/breadth ratio. In addition, its pinnule lobes show a more irregular outline and are less deeply incised.
51 Stratigraphic and geographic distribution.Renaultia footneri is an uncommon species. It ranges from lower Langsettian to upper Bolsovian. The type material is from upper Langsettian strata of the Lancashire coalfield in England. Brousmiche (1983) regarded this species as rare in Saar-Lorraine, where it ranges from upper Westphalian B to middle Westphalian C (upper Duckmantian to middle Bolsovian).
52 Occurrence in the Maritime Provinces, Canada. Cumberland Basin (Nova Scotia): Bell (1944): locality 1052 (GSC 8619 ― together with Corynepteris angustissima and Cordaites sp.).
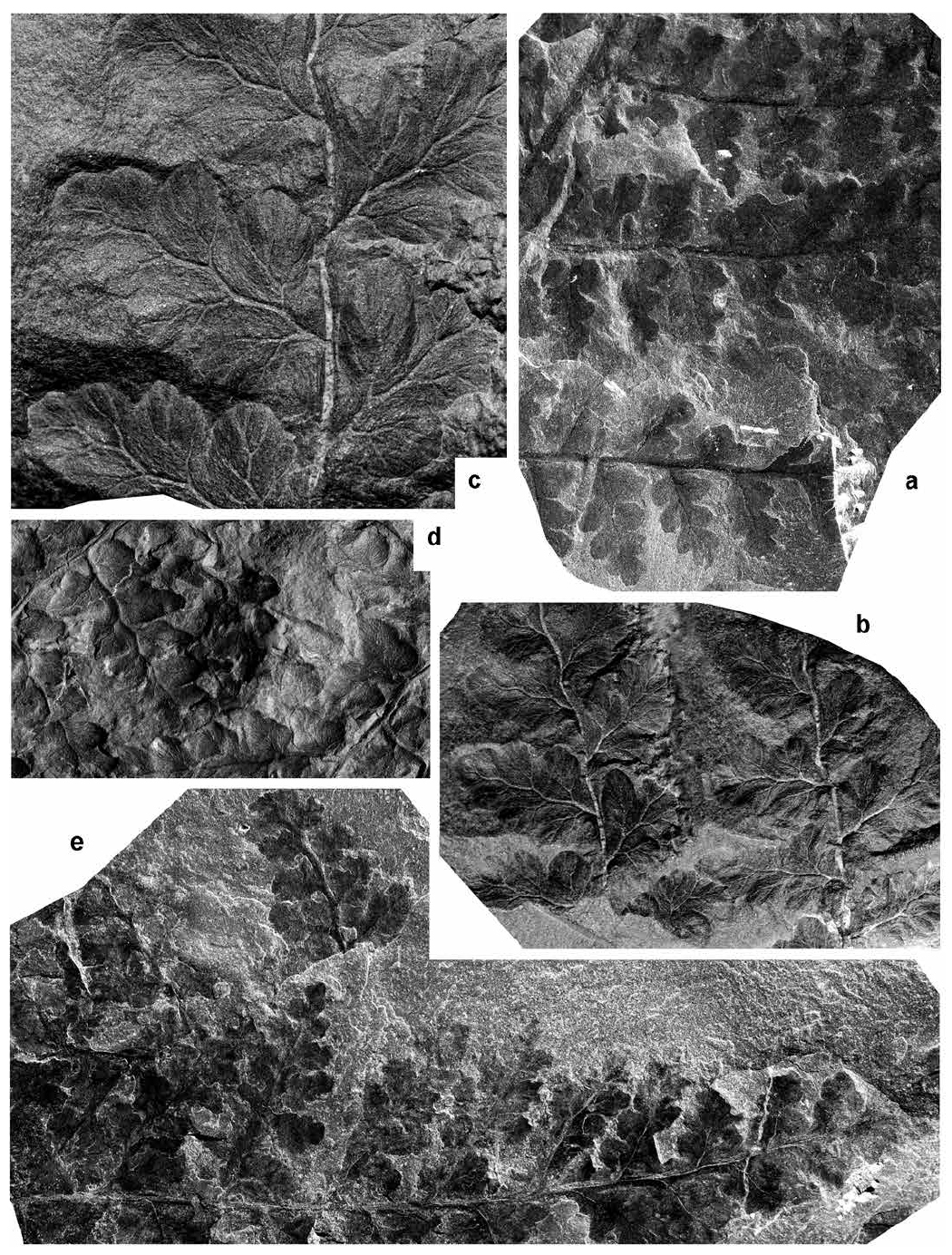
Renaultia rotundifolia (Andrae 1869) Zeiller 1899
(Figs. 7a–c, 8a–e)
 Display large image of Figure 7
Display large image of Figure 7
 Display large image of Figure 8
Display large image of Figure 8
- * ? 1862 Sphenopteris marginata Dawson, p. 321, pl. XV, figs. 38a–b (drawings).
- ? 1868 Sphenopteris marginata Dawson, p. 551, Fig. 192D (same specimen as Dawson 1862).
- * 1869 Sphenopteris rotundifolia Andrae, p. 37, Taf. XII, figs. 1–2a.
- ? 1871 Sphenopteris marginata Dawson, p. 52, pl. XVI, fig. 184.
- ? 1888 Sphenopteris marginata Dawson, p. 73, fig. 23D.
- § 1899 Renaultia rotundifolia (Andrae) Zeiller, p. 17.
- ? 1914 Sphenopteris marginata Dawson (= S. rotundifolia); Stopes, p. 32–33, pl. VIII, fig. 18 (photographic reproduction of Dawson 1871, pl. XVI, fig. 184); pl. VIII, figs. 19–20; text-figs. 1a–b.
- 1923 Renaultia rotundifolia (Andrae) Zeiller; Kidston, p. 317–319, pl. LXXX, figs. 1–6b.
- ? 1937 Sphenopteris Laurenti Andrae; Jongmans, p. 407, pl. 27, figs. 74–76.
- v 1944 Sphenopteris (Renaultia) rotundifolia (Andrae) Zeiller; Bell, p. 71, pl. XIX; pl. XX, fig. 5 (herein Fig. 7a).
- * v 1944 Renaultia hydei Bell, p. 72, pl. XX, fig. 4; pl. XXI, figs. 2–6; pl. XXII, fig. 6 (holotype); pl. XXVI, fig. 6.
- v 1944 Sphenopteris mixta Schimper; Bell, p. 64, pl. VIII, figs. 1, 3 (see Fig. 8d).
- v 1944 Sphenopteris stipulataeformis (Stur) Gothan; Bell, p. 66, pl. XIV, fig. 6 (herein Figs. 7b–c).
- 1966 Renaultia rotundifolia (Andrae) Zeiller; Bell, p. 14, pl. VI, fig. 3.
- 1980 Sphenopteris cf. hoeninghausi Brongniart; Zodrow and McCandlish, p. 67, pl. 90, figs. 1–2.
- T 1983 Renaultia rotundifolia (Andrae) Zeiller; Brousmiche, p. 173–178 (including synonymy), pl. 43, figs. 2–2a (same as Stur 1885, Taf. XLIV, figs. 1, 3), fig. 3 (same as Sphenopteris laurentii, Kidston 1923, pl. IX, fig. 1), figs. 4–5a (same as Kidston 1923, pl. LXXX, figs. 2–3), figs. 6–6a (same as Zeiller 1886, pl. IX, fig. 4), figs. 7–7a, figs. 8–8a (neotype), figs. 9–10a; text-figs. 50–51.
- v 2010 Renaultia rotundifolia (Andrae) Zeiller; Wagner and Álvarez-Vázquez, p. 257, 266, 268.
- 2016 Renaultia rotundifolia (Andrae) Zeiller; Opluštil et al., p. 172, fig. 15F.
53 Description. Frond at least quadripinnate. Broadest visible rachis straight, flat, longitudinally striate, ca. 7 mm wide. Antepenultimate rachis perpendicularly inserted, straight, flat, longitudinally striate, 2–3 mm wide. Pinnae of the penultimate order alternate, apparently lanceolate (incomplete), laterally touching or slightly overlapping. Dimensions: up to 70 mm long and 30 mm broad. Penultimate rachis perpendicularly inserted, rounded, straight or slightly flexuous, smooth, slender, up to 0.75 mm wide. Last order pinnae alternate, lanceolate to subtriangular, closely spaced but not touching laterally. Dimensions: 15–40 mm long and 5–10 mm broad; length/breadth ratio = 3–4. Last order rachis inserted at 50–85°, rounded, straight or slightly flexuous, smooth, winged due to fusion of decurrent pinnule bases, slender, up to 0.50 mm wide. Pinnules alternate, closely spaced, with a relatively thick, convex lamina and undulate margins showing an irregular outline. Less-developed pinnules confluent, ovate, almost entire or very slightly lobed, with a minor contraction on the anadromous side. Dimensions: up to 3 mm long and 1.6 mm broad; length/breadth ratio = 1.7–1.8. Fully developed pinnules shortly stalked, subtriangular, with a symmetrical development of rounded lateral lobes separated by very shallow sinuses. The transition from pinnule to last order pinnae occurs when a fourth pair of lobes is developed. Dimensions: up to 10 mm long and 6 mm broad; length/breadth ratio = 1.6. Veins deeply embedded in the convex lamina. Midrib thin, slightly flexuous and well developed up to near the pinnule apex. Thin lateral veins diverge to each lobe; lateral veins are once to three times dichotomized at a wide angle. Fertile pinnules resembling the vegetative ones, with small, oval and near-marginal sporangia.
54 Remarks. Bell (1944) figured and described Renaultia rotundifolia on the basis of large pinna fragments displaying thin, rounded rachises and shortly stalked, subtriangular pinnules with rounded lobes; these are fully characteristic of the species. Brousmiche (1983), who described Renaultia rotundifolia in detail and reviewed the literature, accepted Bell’s (1944) record.
55 Bell (1944) introduced a new species, Renaultia hydei, which he compared with Renaultia rotundifolia, but distinguished by its smaller pinnules with a less rounded outline. Bell’s specimens are fragmentary and poorly preserved, but I have re-examined the type material of Renaultia hydei and see no evidence for separating this species from Renaultia rotundifolia. One of Bell’s specimens (GSC 9374 ― Bell 1944, pl. XXI, fig. 3) shows rather small, marginal sporangia that, although poorly preserved, confirm the attribution to Renaultia.
56 I also include in Renaultia rotundifolia the specimen from Springhill figured as Sphenopteris stipulataeformis by Bell (1944, pl. XIV, fig. 6; see Figs. 7b–c herein). Sphenopteris stipulataeformis is a species based on a single specimen called Oligocarpia? stipulataeformis by Stur (1885, Taf. XXXIII, fig. 5). I regard Stur’s species as a pteridosperm, synonymous with Karinopteris obtusifolia.
57 Finally, the specimen figured as Sphenopteris mixta by Bell (1944, pl. VIII, figs. 1, 3) was excluded by Brousmiche (1983) from her synonymy of Oligocarpia mixta, albeit without comment. This specimen shows closely spaced, shortly stalked, subtriangular, symmetrical pinnules with rounded, entire margins and a convex lamina (see detail in Fig. 8c). It is included here in Renaultia rotundifolia.
58 It is important to note that most authors, including Bell, accepted Stopes’s (1914) opinion that Renaultia rotundifolia is synonymous with Dawson’s (1862) poorly defined Sphenopteris marginata. In this case, Sphenopteris marginata would be the name that would have priority. However, the poor quality of Dawson’s (1862, 1868) schematic illustrations, the small, fragmentary nature of the specimens, and the incomplete description (Dawson 1871) have convinced most authors (e.g., Kidston 1923; Bell 1944; Brousmiche 1983) that Andrae’s (1869) name, Renaultia rotundifolia, should be preferred, as it is based on more adequate illustrations and more complete description. Only Stopes (1914), who refigured one of Dawson’s specimens (Stopes 1914, pl. VIII, fig. 18) and illustrated two additional specimens (her pl. VIII, figs. 19–20) from the Fern Ledges locality at Saint John, insisted on the priority of Dawson’s name.
59 As in the case of Sphenopteris pilosa, I have not been able to review the holotype of Sphenopteris marginata. I have included Dawson’s Sphenopteris marginata in the list of synonymy with doubts, waiting the opportunity to review the type. If following review the identity of Sphenopteris marginata and Renaultia rotundifolia is confirmed, the conservation of the name Renaultia rotundifolia should be proposed.
60 Comparisons. Fully developed pinnules of Renaultia footneri also possess subtriangular pinnules composed of three or four pairs of rounded lobes; however, these are smaller and possess a higher length/breadth ratio than in Renaultia rotundifolia. Renaultia gracilis has smaller, subrectangular pinnules with a straight midrib and up to three pairs of lobes which are separated by deeper sinuses. At the same stage of development, Sphenopteris rutaefolia possesses larger pinnules with a more asymmetrical aspect. Fully developed pinnules are oblong to oblong-lanceolate and composed of three pairs of lobes separated by shallow sinuses. They also show a clearly developed basal lobe that differs from the more symmetric pinnules of Renaultia rotundifolia.
61 Stratigraphic and geographic distribution.Renaultia rotundifolia is relatively widespread, but never common. The neotype is from the upper Langsettian of the Ruhr District in western Germany. Its total range is from upper Langsettian to lower Asturian, but it seems to characterize Bolsovian strata (Brousmiche 1983).
62 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Bell (1944): locality 77 (GSC 5579 + GSC 9374 ― paratypes of Renaultia hydei); locality 999 (GSC 8610 + GSC 8611 ― part and counterpart + one piece without catalogue number ― together with Sphenophyllum cuneifolium, Calamites sp., Asterophyllites sp. and Lepidostrobophyllum sp.); locality 1070 (GSC 10174 ― figured as Sphenopteris stipulataeformis); locality 1082 (GSC 9368); locality 1086 (GSC 10070); locality 1401 (GSC 9382 ― as Renaultia hydei); locality 1406 (GSC 5547 ― holotype of Renaultia hydei + GSC 9068 ― as Renaultia hydei); locality 1435 (GSC 5571 ― as Renaultia hydei); locality 3040 (GSC 5983 ― together with Cyperites sp.; figured as Sphenopteris mixta); locality 3115 (two pieces without catalogue number ― together with Sphenophyllum cuneifolium and Bowmanites cuneifolius). Donald Reid collection, Joggins, Nova Scotia (1999): DRC–997 55 (together with Cordaites principalis) + DRC–997 56. Saint John (New Brunswick): Dawson (1862, 1868, 1871, 1888). Stopes (1914). Bell (1944): locality 352 (GSC 119c + GSC 111l ― together with Laveineopteris polymorpha + GSC 15033 + one piece without catalogue number ― all labelled as Sphenopteris marginata); locality 804 (GSC 239 ― together with Laveineopteris polymorpha + one piece without catalogue number); locality 2254 = 804 (nine pieces without catalogue number ― together with Laveineopteris polymorpha). New Brunswick Museum collection: Fern Ledges at Saint John, New Brunswick: NBMG 2312 + NBMG 10529 (labelled as Sphenopteris marginata) + NBMG 12046/1; NBMG 12054 (labelled as Sphenopteris marginata). Sydney Basin (Nova Scotia): Zodrow and McCandlish (1980).
63 Occurrence in the United States. West Virginia: Jongmans (1937).
Renaultia schatzlarensis (Stur 1885) Kidston 1890
(Figs. 9a–c)
 Display large image of Figure 9
Display large image of Figure 9
- * p 1885 Hapalopteris Schatzlarensis Stur, p. 58–63, Taf. XXXIX, figs. 7–7a; Taf. XL, figs. 3–6; non Taf. XL, figs. 1–2 (= Boweria schatzlarensis acc. to Frojdová et al. 2017b).
- * 1885 Hapalopteris Aschenborni Stur, p. 63–64, Taf. XXXIX, fig. 6; text-figs. 12a–b (acc. to Gothan 1913).
- § 1890 Renaultia schatzlarensis (Stur) Kidston, p. 32.
- * 1914 Sphenopteris deltiformis Kidston, p. 85–86, pl. X, figs. 9–9a.
- 1923 Sphenopteris (?Renaultia) Schatzlarensis (Stur) Kidston, p. 123–125, pl. XXIX, fig. 4; pl. XXX, figs. 1–1a.
- 1923 Sphenopteris deltiformis Kidston, p. 106–107, pl. XXII, figs. 2–2a (same as Kidston 1914, pl. X, figs. 9–9a).
- p 1938 Sphenopteris cantiana Kidston; Bell, pl. VIII, figs. 2, 3; non p. 25, pl. IX, figs. 1–2 (resembles Eusphenopteris sauveurii).
- v 1944 Sphenopteris deltiformis Kidston; Bell, p. 67–68, pl. XV, fig. 2; pl. XVII, fig. 4.
- 1956 Renaultia schatzlarensis (Stur) Kidston; Danzé, p. 175–181, pl. XXVI, figs. 1–5; text-figs. 9a–c.
- v ? 1966 Sphenopteris deltiformis Kidston; Bell, p. 12, pl. V, fig. 19 (see Remarks).
- v 1966 Sphenopteris (Renaultia) schatzlarensis (Stur) Kidston; Bell, p. 24, pl. XI, fig. 4.
- ? 1978 Sphenopteris sp.; Gillespie et al., p. 115, 132, pl. 51, fig. 6.
- T 1983 Renaultia schatzlarensis (Stur) Kidston; Brousmiche, p. 179–190 (including synonymy), pl. 44, figs. 1–1a (same as Stur 1885, Taf. XL, fig. 4 — lectotype), figs. 2–2c (same as Stur 1885, Taf. XXXIX, fig. 6), figs. 3–4a, figs. 5–5a (same as Sternberg 1821, Taf. XXVI, fig. 5); pl. 45, figs. 1–1b (same as Gothan 1941, Taf. 56, figs. 1–2a), fig. 2, figs. 3–3a (same as Gothan 1913, Taf. 29, fig. 1), figs. 4, 4b (same as Remy and Remy 1959, text-fig. 98), figs. 5–5a, figs. 6–6a (same as Álvarez Ramis et al. 1978, Taf. 11, fig. 5); pl. 46, text-fig. 52C–I2; text-fig. 53.
- p 1985 Zeilleria delicatula Kidston; Gillespie and Craw-ford, p. 252, pl. II, fig. 3; non pl. II, fig. 4 (= cf. Zeilleria frenzlii), fig. 5 (= Zeilleria frenzlii).
- 1989 Zeilleria delicatula Kidston; Gillespie et al., p. 5, 11, pl. 13, fig. 2 (same as Gillespie and Crawford 1985, pl. II, fig. 3).
- T 1997 Zeilleria delicatula Kidston; Kvaček and Straková, p. 62, pl. 19, fig. 1 (photograph of holotype).
- p 2002 Zeilleria frenzlii (Stur) Kidston; Blake et al., p. 268, 291, pl. XVII, fig. 3; non pl. XVII, fig. 2 (= Zeilleria frenzlii).
- v 2010 Renaultia schatzlarensis (Stur) Kidston; Wagner and Álvarez-Vázquez, p. 257, 266.
- 2011 Sphenopteris delicatula Sternberg; Pšenička & Opluštil, p. 79–81, pl. X, figs. 5–7; text-figs. 8, 9.
- Excludenda:
- v 1944 Sphenopteris (Renaultia?) schatzlarensis (Stur) Kidston; Bell, p. 67, pl. XV, figs. 1, 3 (= Sturia amoena).
- 1967 Sphenopteris schatzlarensis (Stur) Zeiller; Tidwell, p. 29–30, pl. 5, fig. 1 (= Sphenopteris sp. indet. — inadequate illustration, but pinnules are smaller and lobes are more filiform and rounded at the top) (together with Asterophyllites grandis); text-fig. 3C.
- 1975 Sphenopteris schatzlarensis (Stur) Zeiller; Tidwell, pl. 22, fig. 6 (enlargement of Tidwell 1967, pl. 5, fig. 1; see also pl. 24, fig. 6 of the 1998 2nd edition of his book).
- 1966 Sphenopteris sp.; Gillespie et al., p. 104, pl. 32, fig. 3 (difficult to judge from the illustration at natural size, but probably Sturia amoena).
- 1978 Sphenopteris schatzlarensis (Stur) Zeiller; Gillespie et al., p. 109, 115, 132, pl. 51, fig. 1 (same as Gillespie et al. 1966, pl. 32, fig. 3).
- 1981 Sphenopteris (?) schatzlarensis (Stur) Zeiller; DiMichele and Dolph, p. 5, pl. 1, Fig. 7 (possibly Crossotheca crepinii).
- ? 1996 Sphenopteris cf. S. schatzlarensis (Stur) Zeiller; Cross et al., p. 456, fig. 23–19.1 (difficult to judge from the illustration at less than natural size, but maybe Sturia amoena).
- 2002 Sphenopteris schatzlarensis (Stur) Zeiller; Blake et al., p. 287, pl. VIII, fig. 2 (probably Urnatopteris herbacea).
- 2002 Sphenopteris (Renaultia) schatzlarensis (Stur) Kidston; Blake et al., p. 293, pl. XIX, fig. 6 (apparently the same specimen as figured on their pl. VIII, fig. 2, but attributed to a different locality).
64 Description. Frond at least quadripinnate. Sturdiest visible rachis straight, up to 2.5 mm wide, longitudinally striate. Pinnae of antepenultimate order alternate, apparently lanceolate (but incomplete). Antepenultimate rachis inserted at 80–90°, rounded, slender, ca. 0.3 mm wide, slightly flexuous, apparently smooth but longitudinally striate when a carbonaceous lamina is preserved. Pinnae of penultimate order alternate, apparently lanceolate (but incomplete). Penultimate rachis inserted at 80–90°, rounded, ca. 0.25 mm wide, slightly flexuous, smooth or longitudinally striate when a carbonaceous lamina is present. Last order pinnae alternate, closely spaced but not touching laterally; lanceolate, gradually tapering into a slender terminal, with up to six pairs of pinnules. Dimensions: up to 15 mm long and 9 mm wide; length/breadth ratio ≈ 1.7. Last order rachis inserted at an acute angle, rounded, slightly flexuous, apparently smooth, ca. 0.10–0.15 mm wide. Pinnules slightly oblique to the rachis, alternate, decurrent, united by a narrow flange in the highest part of pinnae; subtriangular to ovate, with three to five deeply incised, spreading, simple or bifid, fairly wide and obtusely pointed lobes. First basal basiscopic pinnule smaller than the basal acroscopic one; apical pinnule reduced to a simple or bifid segment. Dimensions: 7–13 mm long and 3–9 mm wide; length/breadth ratio = 1.4–2.3. Nervation well-marked in a very thin lamina. Midrib thin, slightly decurrent, sending off individual veins to each lobe.
65 Remarks. Bell (1966, pl. XI, fig. 4) figured at natural size a magnificent specimen of Sphenopteris schatzlarensis from the Joggins section. Previously, Bell (1944, pl. XV, fig. 2; pl. XVII, fig. 4) had described and figured two other specimens as Sphenopteris deltiformis, a rarely cited species that I regard as synonymous with Renaultia schatzlarensis. Bell (1944, p. 68) did compare Sphenopteris deltiformis with Sphenopteris schatzlarensis but retained the two species separately. When introducing Sphenopteris deltiformis, Kidston (1914) figured only fragmentary specimen (the holotype), which he compared with Sphenopteris bella, a species that Brousmiche (1983) considered synonymous with Sturia amoena. The pinnules of Sturia amoena are smaller and more compact than those of Sphenopteris deltiformis; the latter has pinnules that are more deeply incised and have lobes that are slightly swollen around the middle and end in a relatively blunt point (a deltoid shape according to Kidston).
66 It is noted that a specimen figured as Sphenopteris bella by Zeiller (1899, pl. I, fig. 13) from the Langsettian of Zonguldak, northern Anatolia (Turkey), was assigned by Kidston (1923, p. 106) to Sphenopteris deltiformis. However, this specimen was referred to Sturia amoena by Brousmiche (1983), an opinion that I share. Sphenopteris deltiformis was reported from Strádonice in Bohemia, Czech Republic, by Němejc (1937), but not illustrated. Neither Kidston nor Němejc compared Sphenopteris deltiformis with Renaultia schatzlarensis, and Sphenopteris deltiformis was not mentioned by Brousmiche (1983).
67 Bell (1966, pl. V, fig. 19) figured an additional, poorly preserved, specimen from the Fern Ledges at Saint John as Sphenopteris deltiformis. This specimen has rounded rachises bearing small, thin-limbed pinnules with strongly marked veins. Pointed lobes are visible in those places where the outline of pinnules is well preserved. Although this specimen clearly resembles Renaultia schatzlarensis, it also has similarities with Renaultia crepinii. This specimen can only questionably be included in the synonymy of Renaultia schatzlarensis.
68 Of the two specimens from the Sydney Basin, Nova Scotia, that Bell (1938) figured as Sphenopteris cantiana (a synonym of Sphenopteris schwerinii, according to Brousmiche 1983), one (Bell 1938, pl. VIII, figs. 2–3) is included here in Renaultia schatzlarensis. Although Brousmiche (1983) figured and described Renaultia schatzlarensis exhaustively and discussed its synonymy in detail, she did not mention Bell’s record from Nova Scotia.
69 Comparisons. Fully developed pinnules of Sturia amoena are smaller, oval, and composed of three pairs of lobes that are generally simple and separated by shallow sinuses, which gives them a more compact appearance than those of Renaultia schatzlarensis. Fertile remains are, of course, quite different. Fully developed pinnules of Zeilleria delicatula are smaller and characterized by three filiform, simple or bifid, spreading lobes. Pinnules of Boweria schatzlar ensis are smaller, with three to five rounded lobes attached to the rachis at a single, broad point. Fertile specimens show pinnules with a somewhat reduced lamina and a single sporangium placed at or near the tip of the lobes. Sporangia appear globular rather than ovoid as in Renaultia. Sphenopteris schwerinii has vegetative pinnules with up to three pairs of blunt, less clearly individualized lobes. Also, pinnules are smaller, obliquely inserted, with a markedly decurrent base. Pinnae and pinnules of Sphenopteris bronnii are markedly asymmetric. Well-developed apical growth results in pinnules that are more elongate than those of Renaultia schatzlarensis at the same stage of development.
70 Stratigraphic and geographic distribution.Renaultia schatzlarensis is a relatively common and widespread, long-ranging species, from (Yeadonian?) Langsettian to late Asturian. The lectotype is from an indeterminate horizon in the Schatzlar (Žacléř) beds in the Czech part of the Intra-Sudetic Basin, of early to middle Westphalian age. The holotype of Sphenopteris deltiformis is from the lower Duckmantian of Great Britain. Brousmiche (1983) reported Renaultia schatzlarensis from Saar-Lorraine as particularly frequent in middle Bolsovian to lower Asturian strata.
71 Occurrence in the Maritime Provinces, Canada. Cumberland Basin (Nova Scotia): Bell (1944): locality 1039 (GSC 9445 — together with one specimen figured as Pecopteris (Senftenbergia?) sp., but identified here as Oligocarpia brongniartii + four specimens without catalogue number); locality 1056 (GSC 9273 + GSC 9274 ― fragmentary; together with Cyperites sp.); locality 1069 (GSC 9219 ― figured as Sphenopteris deltiformis and cited as locality 1039 + one piece without catalogue number; together with Senftenbergia plumosa); locality 1380 (GSC 10974); locality 2988 (GSC 9220 ― figured as Sphenopteris deltiformis); locality 5865 (GSC 15063 ― with doubt; see Remarks). Bell (1966): locality 2982 (GSC 15036 ― figured as Sphenopteris deltiformis and wrongly cited as coming from locality 352, Saint John, New Brunswick). New Brunswick Museum: NBMG 7237 + NBMG 7239 ― poorly preserved specimens, both labelled as Sphenopteris harttii). Sydney Basin (Nova Scotia): Bell (1938): locality 513 (GSC 1626 ― figured as Sphenopteris cantiana).
72 Occurrence in the United States. Georgia: Gillespie and Crawford (1985), Gillespie et al. (1989), Blake et al. (2002). Ohio: Cross et al. (1996). West Virginia: Gillespie et al. (1966), Gillespie et al. (1978).
Genus Sphyropteris Stur 1883
- 1883 Sphyropteris Stur, p. 23–24 [655–656].
- 1885 Sphyropteris Stur, p. 16–18.
- 1956 Sphyropteris Stur; Danzé, p. 61–62, 261–262.
- 1983 Sphyropteris Stur; Brousmiche, p. 221–222.
73 Type.Sphyropteris crepinii Stur 1883.
74 Remarks.Sphyropteris is easily recognized when its characteristic, transversely oriented sporangiophores, commonly broken off, are preserved; sporangiophores are borne on a narrow expansion at the tip of last order pinnae and pinnules, to which they are placed at right angles. This very rare genus occurs in upper Namurian and lower Westphalian strata. Stur (1883) assigned Sphyropteris to the eusporangiate ferns, which may be correct. However, Kidston (1923) postulated that it could represent the microsporangia of a pteridosperm, an opinion not shared by subsequent authors and not supported by evidence.
Sphyropteris boehnischii Stur 1883
(Figs. 10a–d)
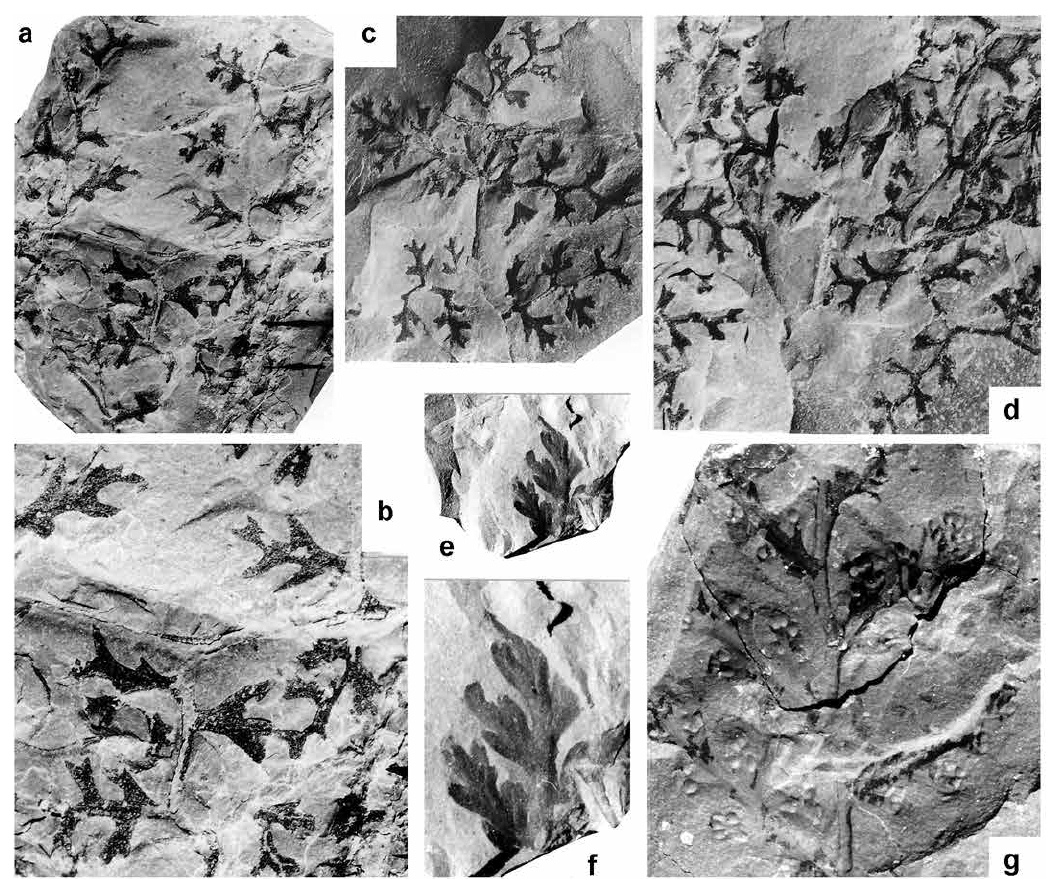 Display large image of Figure 10
Display large image of Figure 10
- 1883 Sphyropteris Boehnischi Stur, p. 24, 26 [656, 658], text-fig. 7 (drawing).
- * 1885 Sphyropteris Boehnischi Stur, p. 17, 24–26, Taf. XXXIX, figs. 3–3a; text-fig. 17 (same as Stur 1883, text-fig. 7).
- * 1912 Sphyropteris frankiana Gothan, p. 246, Taf. 5.
- ? 1913 Sphyropteris cf. Boehnischi Stur; Gothan, p. 132, Taf. 16, fig. 6 (fragmentary).
- 1923 Sphyropteris aff. Frankiana Gothan; Carpentier, p. 4–6, pl. 10, figs. 1–6.
- 1923 Sphyropteris cf. tomentosa Stur; Kidston, p. 365–366, pl. LXXXIV, figs. 8–9; text-fig. 31a–b (the fertile specimen described and figured by Kidston shows widely spaced pinnules subdivided into filiform, bluntly pointed lobes. In contrast, Stur’s Sphyropteris tomentosa possesses ovate pinnules composed of rounded, shorter and wider lobes with shallow sinuses in between).
- 1928 Sphenopteris Boehnischi (Sphyropteris) Stur; Šusta, Taf. XX, fig. 4; Taf. XXIII, fig. 4.
- 1941 Sphenopteris (Sphyropteris) frankiana Gothan; Gothan, p. 22–23, Taf. 57, figs. 1–2a; Taf. 57a, figs. 1–3.
- * v 1944 Rhodea laqueata n. sp. Bell, p. 58, pl. III, fig. 1 (herein Fig. 10d), fig. 2 (herein Fig. 10a), fig. 3 (herein Fig. 10c), figs. 5–6.
- 1957 Sphyropteris Frankiana Gothan; Stopa, p. 67, 184, Tab. XXIII, fig. 7.
- 1966 Rhodea laqueata Bell, p. 12, pl. V, figs. 7–8.
- 1968 Sphyropteris frankiana Gothan; Kotasowa, p. 50, 86, Tab. XXV, fig. 2.
- T 1983 Sphyropteris boehnishii (sic) Stur; Brousmiche, pl. 36, figs. 4–4b (photograph of holotype).
75 Description. Frond at least bipinnate. Penultimate rachis rounded, smooth, slightly zigzagging, ca. 0.5–0.6 mm wide. Last order pinnae well-spaced, probably lanceolate (always incomplete). Last order rachis perpendicularly inserted, rounded, smooth and zigzagging, ca. 0.3–0.4 mm wide. Pinnules widely spaced, inserted at 80–90°; the smaller ones are bifid or reduced to a single, truncate segment; more fully developed ones are composed of three to five bluntly pointed lobes, well spread out, and with an outline parallel to the venation; apical lobe simple or bifid. Dimensions: 3–8 mm long and 1.5–2.5 mm wide. Lamina thick, with a rugose surface. Venation not preserved.
76 Remarks. Bell (1944) figured and described as Rhodea laqueata five fragmentary but distinctive specimens from two localities at Chimney Corner, Cape Breton Island. The loose arrangement of pinnae and the very small, stiff pinnules that are almost devoid of lamina are characteristic. Bell (1944) compared Rhodea laqueata with Rhodea sparsa, another form with small pinnules that are composed of parallel-sided (“linear”) lobes. However, Rhodea sparsa possesses less geniculate axes and has marks of spines on the rachises. This is in keeping with its probable affinity to Lyginopteris hoeninghausii (Kidston 1923, p. 237). Bell (1944) also compared Rhodea laqueata with Boweria minor (now Kidstoniopteris minor ― see Frojdová et al., 2017b), but observed that the pinnule lobes are bluntly pointed in Rhodea laqueata and rounded in Boweria minor.
77 Comparison of Bell’s specimens with more complete and partially fertile material illustrated as Sphyropteris boehnischii by Stur (1885) and Sphenopteris (Sphyropteris) frankiana by Gothan (1941) (the latter being a junior synonym) shows good agreement with regard to pinnae and pinnule morphology and size as well as the stiff “linear” lobes. I consider Sphyropteris boehnischii, Sphyropteris frankiana and Rhodea laqueata to be conspecific.
78 The generic name Rhodea (= Rhodeopteridium Zimmermann, an illegitimate junior homonym) is commonly applied to bipinnate fronds with pinnules characterized by a narrow, generally stiff lamina that follows the outline of repeatedly forked veins. However, pinnules are not as stiff as in Sphyropteris boehnischii. The type of the genus, a common element of Lower Mississippian floras, is Rhodea stachei, a probable progymnosperm, and not a fern like Sphyropteris. Although Bell’s specimens do not preserve the characteristic sporangiate structures, the truncate aspect of some pinnule lobes suggests that the fertile structure has broken off or been shed, suggesting that it belongs to Sphyropteris.
79 Comparisons.Sphyropteris obliqua has smaller pinnules of oval general outline and a wider lamina with two or three pairs of truncate lobes, the uppermost of which is weakly pointed. Vegetative pinnules of Kidstoniopteris minor are smaller and divided into three narrow, parallel-sided, spreading segments terminated by a rounded apex. Its fertile pinnules are totally different, with sporangia arranged in marginally placed groups. Sphenopteris souichii has also vegetative pinnules of uniform size and a narrow lamina following the venation; however, the pinnules are more closely arranged and have rounded lobes and straight, non-flexusous rachises with a fine longitudinal striation. Although Zeiller (1888) compared his Sphenopteris souichii with Sphyro pteris (Sphyropteris boehnischii in particular), no fertile structures of Sphenopteris souichii have yet been recorded.
80 Stratigraphic and geographic distribution.Sphyropteris boehnischii is rarely cited. The holotype is from the lower to middle Westphalian of the Schatzlarer Schichten in the Intra-Sudetic Basin, Czech Republic. Šusta (1928) recorded Sphyropteris boehnischii from Westphalian A and B (Langsettian and Duckmantian) of the Karviner Schichten, in Moravia/Upper Silesia (Czech Republic and Poland). Gothan (1912, 1941) recorded Sphyropteris frankiana from the Westphalian A (Langsettian) of the Ruhr District, western Germany. Stopa (1957) and Kotasowa (1968) figured it from the Westphalian A (Langsettian) of the Upper Silesian Basin in Poland. Carpentier’s (1923) material came from Westphalian B (Duckmantian) strata of Nord/Pas-de-Calais, France.
81 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Bell (1944): locality 1416 (GSC 5673); locality 1420 (GSC 5942 + GSC 5943 ― holotype of Rhodea laqueata + GSC 9366 + GSC 9378). Bell (1966): locality 1420 (GSC 15046).
Family incertae sedis
Genus Boweria Kidston 1911
- 1911 Boweria Kidston, p. 31–34.
- 1923 Boweria Kidston, p. 291–292.
- 1956 Boweria Kidston; Danzé, p. 77–78, 363–365.
- 1986 Boweria Kidston; Brousmiche, p. 232.
- 2017b Boweria Kidston; Frojdová et al., p. 36–47.
82 Type.Boweria schatzlarensis Kidston 1911.
83 Remarks.Boweria is characterized by free circular to oval sporangia that occur in clusters at vein extremities, near to or at the pinnule margin. Fertile pinnules show a slightly reduced lamina in comparision with vegetative ones. The systematic position of Boweria is still uncertain, although it is generally regarded to be a leptosporangiate fern. Kidston (1911) designated Hapalopteris schatzlarensis, from lower Westphalian strata in the Intra-Sudetic Basin (Stur 1885), as type. Boweria is a rarely cited genus. The few available records are all from lower to middle Westphalian strata. The genus comprises only two species: Boweria schatzlarensis and Boweria nowarudensis. Frojdová et al. (2017b) transferred Boweria minor to Kidstoniopteris, a genus with shorter, grouped sporangia and a semi-equatorial annulus.
Boweria schatzlarensis Kidston 1911
(Figs. 10e–g)
- * p 1885 Hapalopteris Schatzlarensis Stur, Taf. XL, fig. 1 (lectotype as designated by Frojdová et al. 2017b), fig. 2; non p. 58–63, Taf. XXXIX, figs. 7–7a (= Renaultia schatzlarensis); Taf. XL, figs. 3–6 (= Renaultia schatzlarensis); text-fig. 11 (= Renaultia schatzlarensis) (acc. to Brousmiche 1983).
- 1911 Boweria Schatzlarensis Kidston, p. 31–34, Fig. 5; Fig. 6A–C (drawings of sporangia).
- 1923 Boweria Schatzlarensis Kidston, p. 292–294, pl. LXXI, figs. 1–6.
- ? p 1938 Boweria schatzlarensis Kidston; Bell, p. 40–41, pl. XXVI, figs. 2–3 (difficult to judge from the illustrations); non pl. XXVIII, figs. 1–2 (difficult to judge, but resembling Renaultia).
- v 1944 Boweria schatzlarensis Kidston; Bell, p. 71, pl. XVIII, fig. 3 (herein Figs. 10e–f); pl. XXI, fig. 1 (herein Fig. 10g).
- 1956 Boweria schatzlarensis Kidston; Danzé, p. 365–370, pl. LX, figs. 1–3a; text-figs. 32a–d.
- 1986 Boweria schatzlarensis Kidston; Brousmiche, p. 232–233, pl. I, figs. 1–1a (same as Kidston 1923, pl. LXXI, figs. 3, 4), fig. 1b (sporangia), figs. 1c–1g (spores); pl. I, figs. 3–5; pl. 1, figs. 5a–5c (spores).
- 1991 Sphenopteris (Boweria) schatzlarensis (Kidston) Jongmans; Josten, p. 214–215, Taf. 97, figs. 3–3a; Abb. 134.
- T k 2017b Boweria schatzlarensis Kidston; Frojdová et al., p. 37–43, pl. I, figs. 1–2 (lectotype), figs. 3–7; pl. II, fig. 1, figs. 2–7 (cuticles); pl. III, figs. 1–4; pl. IV, figs. 1–5; pl. V, figs. 1–8 (sporangia); pl. VI, figs. 1–4 (spores); text-figs. 4–9 (drawings).
84 Description and remarks. Bell (1944) figured as Boweria schatzlarensis two small fragments, one representing vegetative remains and the other fertile remains. Both specimens are well illustrated and come from locality 1386, in the upper part of the Joggins section in Nova Scotia. The vegetative fragment (Bell 1944, pl. XVIII, fig. 3 — Figs. 10e–f herein) shows only six incomplete pinnules in near-terminal position in a last order pinna. These pinnules are obliquely inserted, decurrent, and attached to the rachis at a single, rather broad point; they consist of three to five rounded lobes. The fertile specimen (Bell 1944, pl. XXI, fig. 1 — Fig. 10g herein) shows pinnules with a somewhat reduced lamina and apparently single sporangia placed in a marginal position. This accords well with the sporangia of Boweria schatzlarensis figured by Kidston (1923, pl. LXXI, fig. 3a), Brousmiche (1986, pl. I, figs. 1–1a) and Frojdová et al. (2017b, pl. IV, figs. 2, 3). Sporangia appear globular rather than ovoid, which suggests affinity with Boweria rather than with Renaultia. However, the absence of a visible annulus makes a clear assignment problematic. Sporangia of Boweria schatzlarensis are characterized by a prominent annulus, whereas Renaultia has exannulate sporangia. Kidston (1923) recorded the presence of a single sporangium near the tip of each pinnule lobe.
85 Comparisons.Boweria nowarudensis has smaller, more deeply incised, lobate pinnules of elongate ovoid shape, with five to seven lobes showing bluntly rounded sub-lobes. Vegetative pinnules of Kidstoniopteris minor are smaller and divided into more filiform, rounded or slightly pointed, spreading segments; its fertile pinnules are practically devoid of lamina, with the short-stalked sporangia either solitary or irregularly grouped on the pinnule margin (Frojdová et al., 2017b).
86 Stratigraphic and geographic distribution.Boweria schatzlarensis has been recorded only occasionally. According to Frojdová et al. (2017b) the type material is from Langsettian–Duckmantian strata (Lower Moscovian) of the Ruhr Basin, western Germany. Josten (1991) noted that this species occurs sporadically in the upper Westphalian A and Westphalian B (upper Langsettian and Duckmantian) of western Germany. It has been recorded from Duckmantian strata of Great Britain by Kidston (1923). Danzé (1956) and Brousmiche (1986) noted that its range extended into lower Westphalian C (lower Bolsovian) strata.
87 Occurrence in the Maritime Provinces, Canada. Cumberland Basin (Nova Scotia): Bell (1944): locality 1386 (GSC 5592 + GSC 11006). Sydney Basin (Nova Scotia): Bell (1938): locality 727 (GSC 2924 ― with doubt); locality 989 (GSC 8398 ― with doubt).
Genus Germera Brousmiche 1983
- 1983 Germera Brousmiche, p. 225–226.
88 Type.Germera mendescorreae (Teixeira 1939) Brousmiche 1983.
89 Remarks. The genus Germera has fertile and vegetative pinnules that are identical to each other in general aspect. Fertile pinnules are characterized by eusporangiate sori placed in apical position; sporangia are short, ovoid, 20–26 in number, and inserted on an oblong receptacle, in pendent position below the lamina. Spores are unknown.
Germera brousmicheae sp. nov.
(Fig. 11c)
 Display large image of Figure 11
Display large image of Figure 11
- * 1975 Sphenopteris digitata Doubinger; Doubinger and Germer, p. 375, pl. I, fig. 3 (wrongly cited as fig. 2).
- 1978 Sphenopteris (Zeilleria) delicatula Kidston; Álvarez-Ramis et al., p. 17, Taf. 8, figs. 1–2; Abb. 27 (included in Germera mendescorreae by Brousmiche 1983).
- T 1978 Sphenopteris digitata Doubinger; Álvarez- Ramis et al., p. 17–18, Taf. 8, figs. 3–4 (same as Doubinger and Germer 1975); Abb. 28.
- T p 1983 Germera mendescorreae (Teixeira) Brousmiche, p. 227–233, pl. 55, figs. 1–1e (same as Doubinger and Germer 1975, pl. I, fig. 3, and Álvarez-Ramis et al. 1978, Taf. 8, fig. 3), figs. 2, 4–5; pl. 56, figs. 1–1a, figs. 2–2a (same as Álvarez-Ramis et al. 1978, Taf. 8, figs. 1–2), figs. 3–7c; non pl. 55, figs. 3–3b (topotype of Sphenopteris mendescorreae).
- v 1995 Germera cf. mendescorreae (Teixeira) Brousmiche; Álvarez-Vázquez, p. 181–183, lám. 61, figs. 1–5.
- v 1999 Germera cf. mendescorreae (Teixeira) Brousmiche; Álvarez-Vázquez, p. 44, fig. 4 (name only).
- v 2010 Germera cf. mendescorreae (Teixeira) Brousmiche; Wagner and Álvarez-Vázquez, p. 266, 316.
90 Holotype. Specimen figured and described as Sphenopteris digitata in Doubinger and Germer (1975, p. 375, pl. I, fig. 3); Álvarez Ramis et al. (1978, p. 17–18, Taf. 8, figs. 3–4, Abb. 28); and refigured as Germera mendescorreae by Brousmiche (1983, pl. 55, figs. 1–1e). Collection: Saarbergwerke in Saarbrücken, western Germany, nº C/7440.
91 Derivation of name. After Claudine Brousmiche, in recognition of her significant contribution to the knowledge of sphenopterid ferns of the Carboniferous.
92 Type locality, horizon and age. Hailigenwald Formation, Saarbrucken Group, Saarland, western Germany; Bolsovian–early Asturian.
93 Diagnosis. Less developed pinnules tongue-shaped, decurrent on catadromous side; alternate differentation of lobes and apical development predominant; lobes sturdy, with rounded or tongue-shaped apices; pinnules asymmetric due to early division of basal anadromous lobe; catadromous basal lobe generally simple; pinnules fully developed at the stage of development of the fourth pair of lobes. Fertile pinnules of identical shape to vegetative ones, with fertile structure of Germera-type. (Shortened from the freely translated French decription of the material cited as Germera mendescorreae by Brousmiche 1983, p. 227.)
94 Description. Frond at least bipinnate. Pinnae of the antepenultimate order closely spaced, apparently lanceolate (always incomplete). Dimensions: up to 40 mm long and 22 mm wide. Last order rachis straight, rounded, finely striate longitudinally, ca. 0.6–0.7 mm wide. Last order pinnae alternate, closely spaced and touching at their margins; subtriangular, slightly asymmetric, and tapering gradually into a small, filiform, apical segment that is partially fused with the nearest contiguous pinnules. Dimensions: 10–15 mm long and 8–9 mm wide; length/breadth ratio = 1.25–1.65. Last order rachis straight, flat, winged, finely striate longitudinally, ca. 0.4–0.5 mm wide. Pinnules alternate, closely spaced, inserted at 45–50º, and consisting of up to three pairs of closely adpressed, parallel-sided, elongate lobes; pinnules show a decurrent base and a rounded apex. Dimensions: 4–9 mm long and 1.5–3 mm wide; length/breadth ratio = 2.6–3. Lamina thin, of delicate appearance. Venation consists of a thin midrib with lateral veins at a wide angle, sending off one lateral vein to each lobe.
95 Remarks. This species, here recorded for the first time from Canada, is represented by three specimens from Spicer’s Cove (collection of Fundy Geological Museum, Parrsboro).
96 The generic name Germera was established by Brousmiche (1983, p. 225) for the reproductive structures of a partially fertile specimen that had been illustrated under the name Sphenopteris digitata by Doubinger and Germer (1975, pl. I, fig. 3) and Álvarez-Ramis et al. (1978, Taf. 8, figs. 3–4). Brousmiche (1983) included this specimen, which comes from upper Westphalian strata of Saar-Lorraine, in Germera mendescorreae. Subsequently, specimens from lower Bolsovian strata of the Peñarroya-Belmez-Espiel coalfield, southwestern Spain, were recorded as Germera cf. mendescorreae by Álvarez-Vázquez (1995, 1999) and Wagner and Álvarez-Vázquez (2010).
97 The species name Sphenopteris digitata, as first used by Doubinger (1959, p. 114), was published without a diagnosis or illustrations. Vetter (1968, pl. XL, figs. 5–6), Álvarez-Ramis and Doubinger (1973, pl. 1, figs. 1–1a), and Boureau and Doubinger (1975, figs. 443A, B) figured the cited specimen at a later date. This specimen came from Stephanian strata in the Saint-Perdoux Basin, Lot, southern France. It is a small fragment of penultimate order pinna showing small pinnules with two or three pairs of short, filiform lobes, a decurrent base and a rounded apex.
98 Brousmiche (1983) accepted both that the Westphalian material of Germera from Saar-Lorraine was conspecific with the Portuguese species Sphenopteris mendescorreae, and the synonymy between Teixeira’s species Sphenopteris mendescorreae and Sphenopteris sampaiana, as proposed by Wagner and Lemos de Sousa (1983). Both Sphenopteris mendescorreae and Sphenopteris sampaiana were described by Teixeira (1939) from Stephanian C strata in the Douro Basin, northern Portugal. Álvarez-Vázquez (1995) refigured a number of syntypes of Sphenopteris mendescorreae and Sphenopteris sampaiana and noted that pinnules of Sphenopteris sampaiana retain the same size throughout the pinna and are characterized by stiff, simple or bifid lobes with acute apices (see Fig. 11a herein). In contrast, Germera mendescorreae (originally Sphenopteris mendescorreae) possesses more spreading pinnules, constituted by filiform, parallel-sided lobes with more rounded apices (Fig. 11b). Germera mendescorreae and Sphenopteris sampaiana are thus separate species.
99 As pointed out by Álvarez-Vázquez (1995) and Wagner and Álvarez-Vázquez (2010, p. 316), the Westphalian specimens from Saar-Lorraine attributed to Germera mendescorreae by Brousmiche (1983) actually represent a different species of Germera, described here as Germera brousmicheae sp. nov.
100 Comparisons.Germera mendescorreae has pinnules that are more spaced-out and of smaller size and lower length/ breadth ratio than those of Germera brousmicheae. Fully developed, vegetative pinnules of Zeilleria frenzlii, although similar in size to those of Germera brousmicheae, possess larger, more spread-out lobes with acute apices. Also, the reproductive structures of Zeilleria differ from those of Germera. Pinnules of Sphenopteris sampaiana retain the same size throughout the pinna and possess stiff, simple or bifid lobes with acute apices
101 Stratigraphic and geographic distribution.Germera brousmicheae is known only from a few localities, insufficient to establish a stratigraphic range. The specimens described by Brousmiche (1983) from Saar-Lorraine are from upper Bolsovian to upper Asturian strata. Álvarez-Vázquez (1995, 1999) and Wagner and Álvarez-Vázquez (2010) recorded material from lower Bolsovian strata in the Peñarroya Basin of southwestern Spain.
102 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Fundy Geological Museum: FGM 999 GF 13.8/21.2 + FGM 999 GF 13.9/21.2 (together with Dorycordaites palmaeformis and rootlets) + FGM 997 GF 31.7 (fragmentary ― together with Dorycordaites palmaeformis).
Order Filicales Copeland 1947
Family Sermayaceae Eggert and Delevoryas 1967
Genus Oligocarpia Göppert 1841
- 1841 Oligocarpia Göppert, p. 35.
- 1883 Oligocarpia Göppert; Stur, p. 54 [686].
- 1954 Oligocarpia Göppert; Abbott, p. 40–44.
- 1956 Oligocarpia Göppert; Danzé, p. 48–50, 289–292.
- 1983 Oligocarpia Göppert; Brousmiche, p. 234–235.
103 Type.Oligocarpia gutbieri Göppert 1841.
104 Remarks. Fertile pinnules of Oligocarpia are characterized by circular sori attached to the abaxial side of pinnules; each sorus is composed of three to thirty subspherical to pyriform, stalked sporangia showing a prominent annulus consisting of two rows of thick-walled cells. Individual sori cover lateral veins; most often these are sufficiently large to occupy the entire space between midrib and margin of the (slightly reduced) pinnule lamina; if not they are situated near the margin. Vegetative pinnules are either sphenopteroid or pecopteroid, and similar in shape to the fertile ones, with similar or a slightly smaller size. Oligocarpia is a relatively common genus with several species. It occurs widely in Westphalian as well as Stephanian strata.
Oligocarpia brongniartii Stur 1883
(Figs. 12a–b)
 Display large image of Figure 12
Display large image of Figure 12
- * ? 1871 Sphenopteris splendens Dawson, p. 53, pl. XVI, figs. 186–186a (diagrammatic drawings — see Remarks).
- 1883 Oligocarpia brongniarti Stur, p. 56 [688], Figs. 16a–c (see Stopes 1914).
- * 1885 Oligocarpia Brongniarti Stur, p. 131–134, Taf. LVII, figs. 2–3; text-fig. 20.
- ? 1914 Oligocarpia splendens (Dawson) Stopes, p. 40–41, pl. X, fig. 24 (photograph of holotype); text-fig. 4 (drawing).
- 1938 Oligocarpia brongniarti Stur; Bell, p. 42–43, pl. XXIX; pl. XXX, figs. 1–2.
- v 1944 Oligocarpia brongniarti Stur; Bell, p. 76, pl. XXV, figs. 2, 4 (poorly preserved).
- v 1944 Pecopteris (Senftenbergia?) sp.; Bell, p. 86, pl. XLII, figs. 2, 4 (herein Figs. 12a–b).
- 1954 Oligocarpia brongniarti Stur; Abbott, p. 49–51, figs. 6–12 (drawings), 17–19b (drawings of sporangia and sori).
- 1962 Oligocarpia brongniarti Stur; Bell, p. 15–16, pl. I, fig. 3.
- 1966 Oligocarpia brongniarti Stur; Bell, p. 40, pl. XIX, fig. 4.
- T 1983 Oligocarpia brongniartii Stur; Brousmiche, p. 245–249, pl. 62, figs. 1–4; pl. 63, figs. 1–1f (neotype), fig. 2; pl. 64, figs. 1–3.
105 Description. Frond at least bipinnate. Penultimate rachis straight, rounded, ca. 0.25 mm wide. Last order pinna alternate, lanceolate, inserted at 45–50°. Dimensions: up to 12 mm long and 4 mm wide at the base; length/breadth ratio = 3. Last order rachis straight, rounded, ca. 0.1 mm wide. Pinnules alternate, oblique, ovate, slightly contracted at the base with a narrow sinus or attached by their entire width (pecopteroid aspect); decurrent, with slightly lobate or toothed margins; terminal subcircular, very short. Dimensions: 1–2 mm long and 0.5–1 mm wide; length/breadth ratio = 0.5–2. Lamina relatively thick. Venation well-marked, showing a distinct, relatively wide, flexuous midrib, giving off a single vein to each lobe.
106 Remarks. Stopes (1914) considered Dawson’s Spheno pteris splendens (which she transferred to Oligocarpia) to be con-specific with Stur’s Oligocarpia brongniartii. However, Abbott (1954), who re-examined the holotype of Sphenopteris splendens, concluded that the two species were not synonymous based on different lobing patterns and pinnule margins. Dawson’s material did not include fertile remains, which put the synonymy further into doubt. I follow Bell (1938) and recommend that the name Sphenopteris splendens be restricted to its type material; its synonymy with Oligocarpia brongniartii is open to question. The present material is thus attributed to Oligocarpia brongniartii, which is based on much better, well-described material.
107 Although fragmentary and poorly preserved, the specimen figured as Oligocarpia brongniartii by Bell (1944, pl. XXV, figs. 2, 4) seems a good fit for this species. The same locality at Springhill (locality 1039) yielded a specimen figured by Bell (1944, pl. XLII, figs. 2, 4; Figs. 12a–b herein, after removal of occluding material with a vibratory tool) as Pecopteris (Senftenbergia?) sp. Although equally fragmentary as the specimen described by Bell as Oligocarpia brongniartii, this specimen is better preserved and clearly identifiable as Oligocarpia brongniartii. Bell compared Oligocarpia brongniartii with Senftenbergia pennaeformis, although this now seems irrelevant.
108 Comparisons. Pinnules of Oligocarpia mixta are smaller than those of Oligocarpia brongniartii, oblong, and with more deeply incised lobes. In addition, the sori of Oligocarpia mixta are smaller, comprising only three to five sporangia, in contrast to seven to twelve in Oligocarpia brongniartii. Vegetative pinnules of Oligocarpia gutbieri are larger and more equidimensional than those of Oligocarpia brongniartii. They are ovate to subtriangular, hardly if at all lobate (often only a sinuous margin); where present, lobes in Oligocarpia gutbieri are rounded and separated by shallow sinuses.
109 Stratigraphic and geographic distribution.Oligocarpia brongniartii is a common and widely represented species. Brousmiche (1983) recorded its range in Saar–Lorraine (Germany–France) as Westphalian C (Bolsovian) to Westphalian D (Asturian). Josten (1991) recorded Oligocarpia brongniartii from upper Westphalian B (Duckmantian) to upper Westphalian D (Asturian) in the Ruhr District of western Germany.
110 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Bell (1944): locality 1039 (GSC 9067 + GSC 9445 ― labelled as Pecopteris (Senftenbergia?) sp; together with Renaultia schatzlarensis). Sydney Basin (Nova Scotia): Bell (1938): locality 487 (GSC 1657); locality 490 (GSC 1649); locality 518 (GSC 1593). Bell (1966): locality 465 (GSC 1648).
111 Occurrence in the United States. Ohio: Abbott (1954).
Family uncertain
Genus Sturia Němejc 1934
- 1934 Štúria Němejc, p. 2–3.
- 1956 Sturia Němejc; Danzé, p. 79, 370.
- 1983 Sturia Němejc; Brousmiche, p. 273–274.
- 2017a Sturia Němejc; Frojdová et al., p. 154.
112 Type.Sturia amoena (Stur 1885) Němejc 1934.
113 Remarks. Němejc (1934) introduced Sturia to accommodate Hapalopteris amoena. He did not give a diagnosis but provided a detailed description of the fertile structure. Sturia’s fertile pinnules possess leptosporangiate sporangia, with small, free, sessile, oval sporangia that cluster in variable numbers. Sturia amoena is the only species of the genus yet described.
Sturia amoena (Stur 1885) Němejc 1934
(Figs. 13a–f)
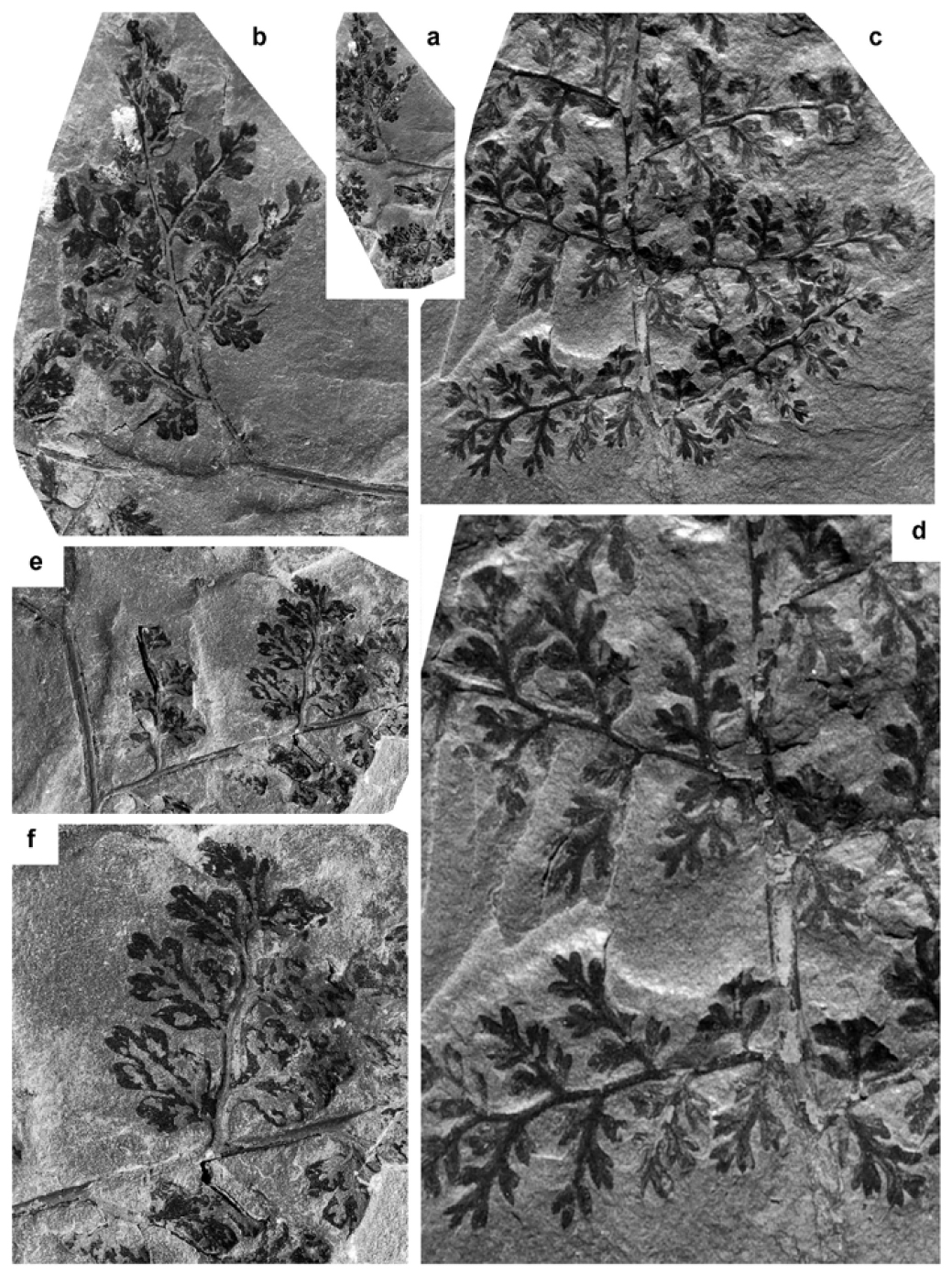 Display large image of Figure 13
Display large image of Figure 13
- * 1885 Hapalopteris amoena Stur, p. 52–54, Taf. XLI, figs. 7–7a.
- * 1885 Hapalopteris bella Stur, p. 49–52, Taf. XLII, figs. 1–2 (acc. to Gothan 1941).
- * 1913 Sphenopteris Schilleri Gothan, p. 147–148, Taf. 30, fig. 2 (acc. to Gothan 1941).
- 1923 Sphenopteris bella (Stur) Zeiller; Kidston, p. 127–129, pl. XXVII, figs. 1–3c.
- * 1923 Sphenopteris quadriloba Kidston, p. 122–123, pl. XXV, figs. 1–2a (two specimens that came from the same horizon as Kidston’s Sphenopteris amoenaeformis).
- * 1923 Sphenopteris amœnæformis Kidston, p. 125– 127, pl. XXV, figs. 6–6a; pl. XXVI, figs. 1–3b (acc. to Danzé 1956).
- § 1934 Štúria amoena Stur (gen. nov.) Němejc, p. 2–3, plate, figs. 1–3, figs. 4–6 (sporangia).
- v 1944 Sphenopteris amoenaeformis? Kidston; Bell, p. 65, pl. XII, figs. 2, 4; pl. XIII, fig. 2.
- v 1944 Sphenopteris (Renaultia?) schatzlarensis (Stur) Kidston; Bell, p. 67, pl. XV, fig. 1, fig. 3 (partly refigured herein as Figs. 13c–f).
- v 1944 Sphenopteris (Zeilleria?) sp.; Bell, p. 74, pl. XXII, fig. 5; pl. XXIII, fig. 1.
- 1956 Sturia amoena (Stur) Němejc; Danzé, p. 370–376, pl. LXXII, figs. 4–4a; pl. LXXIII, figs. 1–1d.
- 1963 Štúria amoena (Stur) Němejc; Němejc, Tab. LII, figs. 1–4, figs. 5–6 (sporangia) (same as Němejc 1934).
- ? 1966 Sphenopteris sp.; Gillespie et al., p. 104, pl. 32, fig. 3 (difficult to judge from the illustration at natural size).
- ? 1978 Sphenopteris schatzlarensis (Stur) Zeiller; Gillespie et al., p. 109, 115, 132, pl. 51, fig. 1 (same as Gillespie et al. 1966, pl. 32, fig. 3).
- T 1983 Sturia amoena (Stur) Němejc; Brousmiche, p. 274–278 (including synonymy), pl. 73, figs. 1–1b (lectotype), figs. 2–2a (same as Stur 1885, Taf. XLII, fig. 1), figs. 3–3a (another part of lectotype), figs. 4–5a (same as Kidston 1923, pl. XXVI, figs. 2–3), figs. 6–6a (same as Kidston 1923, pl. XXV, fig. 6); pl. 74, figs. 1–1a (same as Gothan 1913, Taf. 30, fig. 2), figs. 2–3 (same as Stur 1885, Taf. XLII, fig. 1), figs. 4–8; text-figs. 78A1–79.
- ? 1996 Sphenopteris cf. S. Schatzlarensis (Stur) Zeiller; Cross et al., p. 456, fig. 23–19.1 (figured at less than natural size).
- v 2010 Sturia amoena (Stur) Němejc; Wagner and Álvarez-Vázquez, p. 266, 270.
- T 2017a Sturia amoena (Stur) Němejc; Frojdová et al., p. 154–158, pl. 1, figs. 1–5 (lectotype), figs. 6–10; pl. 2, figs. 1–8 (sporangia and spores); text-figs. 1A–B (drawings).
114 Description. Frond at least tripinnate. Antepenultimate rachis sturdy, straight, flat, smooth, ca. 2 mm wide. Pinnae of penultimate order apparently subtriangular (always incomplete), alternate, spaced out or partly overlapping. Penultimate rachis inserted at 45–50°, slightly flexuous, rounded, smooth, ca. 0.5–0.6 mm wide. Last order pinnae decurrent, alternate, spaced out or touching laterally depending on the position in the frond; subtriangular, with margins gradually tapering to a blunt apical segment. Dimensions: 11–20 mm long and 6–10 mm broad; length/breadth ratio = 1.8–2. Last order rachis inserted at 75–90°, thin, slightly flexuous, rounded, smooth, ca. 0.2 mm wide. Pinnules alternate, oblique to the rachis, well-spaced, thus giving an airy aspect to the pinna. Less developed pinnules obliquely inserted and united to the rachis by only part of the basal width; decurrent, small, subtriangular. Fully developed (and characteristic) pinnules trilobate to quintulobate, almost equidimensional, with slightly divergent, broad and bluntly pointed lobes, separated by shallow sinuses. They pass gradually into pinnae at the development of a fourth pair of lobes. Dimensions: 1.5–3 mm long and 1.5–2.5 mm broad; length/breadth ratio = 1–1.2. Lamina fairly thin to moderately convex. Nervation consists of a thin, decurrent and slightly flexuous midrib that gives off a single vein to each lobe.
115 Remarks. I include in Sturia amoena the two relatively large and fairly well-preserved specimens from Joggins that Bell (1944, pl. XV, figs. 1, 3) figured as “Sphenopteris (Renaultia?) schatzlarensis”. Unfortunately, these specimens have been varnished, obscuring morphological details and making photography difficult. Bell’s illustrations (at natural size) are thus difficult to interpret. Reillustration herein (Figs. 13c–d) of the upper right hand corner of Bell’s pl. XV, fig. 3 nonetheless provides evidence of a strong resemblance between Bell’s material and the holotype of Sturia amoena.
116 Also three poorly figured specimens included in Sphenopteris amoenaeformis? by Bell (1944, pl. XII, figs. 2, 4; pl. XIII, fig. 2 — all from the same locality) are here regarded as belonging to Sturia amoena. Bell’s material undoubtedly resembles Kidston’s (1923) illustrated specimens of Sphenopteris amoenaeformis, a species that was placed in synonymy with Sturia amoena by Danzé (1956), who did not mention Bell’s record.
117 Two moderately well-preserved pinna fragments of the antepenultimate order that Bell (1944, pl. XXII, fig. 5; pl. XXIII, fig. 1) identified as Sphenopteris (Zeilleria?) sp. also seem to accord well with Sturia amoena. Both specimens were re-examined for the present study. Although Bell (1944) compared these specimens with Sturia amoena, he did not include them in that species because he considered the size of penultimate pinnae to be different. The specimen figured by Bell (1944, pl. XXIII, fig. 1) shows a transition from spread-out thin-limbed pinnules to stiffer pinnules with a reduced lamina. The latter are suggestive of fertile pinnules, but no sporangia occur to confirm this possibility. Bell (1944, p. 74) mentioned the presence of poorly preserved sporangia borne distally at the end of lobes, which he thought were split into a four-segmented cupula-like bodies suggestive of Zeilleria. However, my re-examination of the specimen provided no evidence of sporangia. Perhaps, the rough texture of the silty mudstone on which this specimen is imprinted led to Bell’s interpretation.
118 Comparisons. The angle of insertion of pinna and pinnules of Sturia amoena, the small size of pinnules, their subtriangular shape, and the bluntly pointed lobes all suggest a resemblance to Renaultia hemingwayi. However, the latter species shows a basal anadromous pinnule that is relatively longer, thus producing an asymmetry in last order pinnae. This is not present in Sturia amoena. Vegetative pinnules of Renaultia schatzlarensis also possess broad, obtusely pointed lobes, but these are more spread out and more deeply incised. Fully developed pinnules of Sturia amoena are smaller and more equidimensional. The equally small pinnules of Sphenopteris bronnii are longer and composed of up to four pairs of filiform lobes, with acute apices, in contrast to the almost equidimensional, broad and bluntly pointed lobes separated by shallow sinuses in Sturia amoena.
119 Stratigraphic and geographic distribution.Sturia amoena seems to have been misidentified by several authors and so the existing records of its stratigraphic occurrence are suspect. However, it appears that Sturia amoena is relatively widespread and common throughout the Westphalian. The lectotype is from Duckmantian–Bolsovian strata in Belgium. Brousmiche (1983) stated that this species ranges through the Westphalian, and possibly above. She observed its frequent occurrence in lower Westphalian C (Bolsovian) and upper Westphalian D (Asturian) coal-bearing strata of Saar-Lorraine. Frojdová et al. (2017a) figured Duckmantian specimens from the Czech Republic. Gothan’s Sphenopteris schilleri was described from the upper part of the Muldengruppe (i.e. upper Duckmantian) in Upper Silesia, Poland. The type material of Kidston’s Sphenopteris amoenaeformis came from two localities in the Clackmannanshire and Lanarkshire coalfields, Midland Valley of Scotland, from strata of late Langsettian age. The holotype of Sphenopteris quadriloba originated from the same level in the Lanarkshire coalfield. In the Iberian Peninsula, Wagner and Álvarez-Vázquez (2010) recorded Sturia amoena from lower Bolsovian strata of the Peñarroya-Belmez-Espiel coalfield, southwestern Spain, and from the lower Asturian of the Central Asturian Coalfield, northwestern Spain.
120 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Bell (1944): locality 736 (two pieces without catalogue number — together with Dorycordaites palmaeformis and Samaropsis sp.); locality 1125 (GSC 9351 + GSC 9622 — figured as Sphenopteris (Zeilleria?) sp. by Bell); locality 1367 (one piece without catalogue number — poorly preserved); locality 1380 (GSC 10975 + six pieces without catalogue number); locality 1388 (GSC 4449 + GSC 9929 — both figured as Sphenopteris (Renaultia?) schatzlarensis by Bell; together with small fragments of Adiantites adiantoides + GSC 11009 + one piece without catalogue number); locality 1390 (GSC 5576 + GSC 5580 + GSC 9372 — all figured as Sphenopteris amoenaeformis? by Bell + six fragmentary specimens without catalogue number); locality 1416 (two pieces without catalogue number — together with Sphenophyllum cuneifolium); locality 1564 (four pieces; poorly preserved and without catalogue number — together with Sphenophyllum cuneifolium); locality 2986 (one piece without catalogue number). New Brunswick: Bell (1944): locality 655 (two small pieces without catalogue number).
121 Occurrence in the United States. West Virginia: Gillespie et al. (1966); Gillespie et al. (1978).
Order Botryopteridales Meyen 1987
Family Tedeleaceae Eggert and Taylor 1966
Genus Senftenbergia Corda 1845
- 1845 Senftenbergia Corda, p. 91.
- 1938–39 Senftenbergia Corda; Radforth, p. 751–760.
122 Type.Senftenbergia pennaeformis (Brongniart 1822) Stur 1877.
123 Remarks. Vegetative pinnules of Senftenbergia are pecopteroid, united to the rachis by their entire basal width. Fertile pinnules are generally reduced in lamina, and the sporangia normally occupy the entire width between midrib and pinnule margin; they possess annulate, relatively large, free sporangia of pyriform to ovoid shape, placed in lateral position on secondary veins.
Senftenbergia plumosa (Artis 1825) Stur 1877
(Fig. 14)
 Display large image of Figure 14
Display large image of Figure 14
- * 1825 Filicites plumosus Artis, p. 17, pl. XVII.
- * 1832 Sphenopteris caudata Lindley and Hutton, p. 137, pl. 48 (acc. to Kidston 1886).
- * 1834 Sphenopteris crenata Lindley and Hutton, p. 57–58, pls 100–101 (acc. to Kidston 1893).
- * 1834 Pecopteris serra Lindley and Hutton, p. 71–72, pl. 107 (acc. to Kidston 1886).
- * 1836 Pecopteris dentata Brongniart, p. 346–348, pl. 123, figs. 1–5; pl. 124 (acc. to Kidston 1886).
- * 1836 Pecopteris delicatula Brongniart, p. 349–350, pl. 116, figs. 6–6A (acc. to Kidston 1886).
- * 1836 Aspidites Silesiacus Göppert, p. 364–367, Taf. XXVII; Taf. XXXIX, fig. 1 (acc. to Kidston 1886).
- * 1836 Aspidites Glockeri Göppert, p. 375–376, Taf. XXIX, figs. 1–4 (acc. to Kidston 1886).
- * 1838 Pecopteris mucronata Presl in Sternberg, p. 159 (referring to Sternberg 1821, Taf. XVI, fig. 6 – photograph in Kvaček and Straková 1997, pl. 37, fig. 1) (acc. to Němejc 1940).
- * 1854 Pecopteris angustifida Ettingshausen, p. 45, Taf. 16, fig. 1 (acc. to Kidston 1886).
- 1854 Pecopteris Glockeriana Presl in Sternberg; Ettingshausen, p. 44, Taf. 17, fig. 1.
- * 1862 Neuropteris serrulata Dawson, p. 320, pl. XV, figs. 35a–b (diagrammatic drawings — acc. to Stopes 1914).
- 1868 Pecopteris (Alethopteris) serrulata Dawson, p. 534, 548, 553, Fig. 192K (Dawson compared with Pecopteris plumosa, Pecopteris serra, Pecopteris dentata and Pecopteris delicatula).
- * 1868 Pecopteris (Alethopteris) preciosa Hartt in Dawson, p. 534, 548, 553–554, Fig. 192L (although Stopes 1914 refigured these specimens as indeterminable, she compared with Pecopteris plumosa).
- 1871 Pecopteris (Aspidites?) serrulata Dawson, p. 55, 92, pl. XVIII, figs. 207–209 (Dawson compared with Pecopteris silesiaca, a synonym of Senftenbergia plumosa).
- 1871 Pecopteris (Aspidites?) preciosa Dawson, p. 56, 92, pl. XVIII, fig. 210 (specimen figured photographically by Stopes 1914, pl. XXIII, fig. 61, who regarded it as indeterminable), figs. 211– 211a (photographed by Stopes 1914, pl. XXIV, fig. 63).
- 1871 Neuropteris serrulata Dawson, p. 49–50, 92, pl. XVIII, fig. 213 (drawing).
- § 1877 Senftenbergia plumosa (Artis) Stur, p. 187 [293].
- 1879–80 Pecopteris dentata Brongniart; Lesquereux, p. 240–241, pl. XLIV, figs. 4–4a.
- 1879–80 Pecopteris pennaeformis Brongniart; Lesquereux, p. 239–240, Pl. XLV, figs. 1–2a.
- 1896 Dactylotheca plumosa (Artis) Kidston; Kidston, p. 205–219 (including synonymy), pl. I, figs. 1–4b; pl. II, figs. 5–9c, fig. 14 (sporangia); pl. III, figs. 10–13c.
- p 1899 Pecopteris (Dactylotheca) dentata (Brongniart) Zeiller; White, p. 75–78 (including synonymy), pl. XXIV, figs. 1–2; pl. XXV; pl. XXVI, figs. 2–4; pl. XXVII; non pl. XXVI, fig. 1 (resembles Pecopteris micromiltonii).
- 1914 Pecopteris plumosa (Artis) Brongniart; Stopes, p. 44–46, pl. XII, figs. 27–29 (types of Pecopteris serrulata); text-fig. 7 (drawing).
- 1925 Pecopteris pennaeformis Brongniart; Noé, p. 15, pl. XXXII.
- 1925 Pecopteris dentata Brongniart; Noé, p. 15, pl. XXXIII.
- 1937 Dactylotheca plumosa (Artis) Kidston; Jongmans, p. 406, 407, pl. 26, fig. 70.
- k 1937–38 Dactylotheca plumosa (Artis) Kidston; Rad-forth, p. 387–395, pl. I, fig. 1, fig. 2 (sporangium), figs. 3–5, figs. 6–12 (sporangia); pl. II, figs. 1–12 (spores).
- 1938 Dactylotheca plumosa forma dentata Bell, p. 76–77, pl. LXXIII, fig. 3; pl. LXXVII, figs. 3–4; pl. LXXX, figs. 1, 3.
- k 1943 Pecopteris plumosa var. dentata Sterzel; Andrews, p. 432–434, pl. 11, figs. 10–12 (cuticles), figs. 13–14.
- v 1944 Pecopteris (Senftenbergia) plumosa formacrenata Bell, p. 84–85, pl. XXXVIII, figs. 1–4; pl. XXXIX, figs. 5–6.
- ? 1949 Pecopteris plumosa (Artis) Brongniart; Arnold, p. 187, 261, pl. XXVI, fig. 5 (too poorly figured for a proper identification).
- 1951 Pecopteris (Senftenbergia) plumosadentata Dalinval; Corsin, p. 199–208 (including synonymy), pls CXII–CXIV; pl. CXV (as forma crenata); pl. CXVI; pls CXVII, CXVIII (as forma crenata); text-figs. 42a–c (drawing).
- 1957 Dactylotheca plumosa (Artis) Kidston; Janssen, p. 134, 136, Fig. 117q (drawing), Fig. 120.
- 1957 Senftenbergia pennaeformis (Brongniart) Stur; Janssen, p. 138, 139, Fig. 117p (drawing), Fig. 121.
- 1958 Dactylotheca plumosa (Artis) Kidston; Langford, p. 170, Fig. 290; Figs. 291a–c (drawings).
- 1959 Pecopteris (Dactylotheca) plumosa (Artis) Kidston; Canright, p. 31, pl. 3, fig. 15.
- 1959 Asterotheca arborescens (Schlotheim ex Brongniart) Stur; Canright, p. 31, pl. 3, fig. 11.
- 1959 Pecopteris (Senftenbergia) pennaeformis (Brongniart) Stur; Canright, p. 32, pl. 3, fig. 12.
- 1960 Pecopteris (Senftenbergia) plumosadentata Dalinval, p. 51, pls V–XV.
- 1962 Pecopteris (Senftenbergia) pennaeformis (Brongniart) Stur; Bell, p. 35, pl. XXVI, fig. 2.
- v 1966 Senftenbergia plumosa (Artis) Stur; Bell, p. 12, pl. V, fig. 18.
- 1968 Pecopteris dentata Brongniart; Basson, p. 88–89, pl. 17, fig. 2 (poorly figured); pl. 18, fig. 1.
- 1969 Pecopteris plumosa (Artis) Brongniart; Darrah, p. 127–128, 214, pl. 13, fig. 4 (recorded as Pecopteris dentataplumosa in the plate caption).
- 1982 Pecopteris plumosa forma dentata Sterzel; Oleksyshyn, p. 86–88, Figs. 17E–F.
- 2001 Senftenbergia plumosa (Artis) Stur; Bek and Pšenička, p. 218–227, pl. I, figs. 1–3, 6 (spores), figs. 4–5; pl. II, figs. 1–8 (spores); pl. III, figs. 1–3 (sporangia), fig. 4; pl. IV, figs. 1–2 (sporangia), fig. 3; pl. V, figs. 1–2 (sporangia), figs. 3–5 (spores).
- 2002 Pecopteris plumosa (Artis) Brongniart; Blake et al., p. 286, pl. VII, fig. 2.
- k 2003 Senftenbergia plumosa (Artis) Stur; Pšenička and Bek, p. 301, pl. I, figs. 1–3; pl. II, figs. 1–5 (spores); pl. III, figs. 1–5 (cuticle); pl. IV, figs. 1–4 (cuticle); text-figs. 3–6.
- 2005 Senftenbergia plumosa (Artis) Stur; Bash-forth, p. 48–51, pl. 11, figs. 1–5; pl. 12, figs. 6, 8; text–fig. 24E.
- 2006 Senftenbergia plumosa (Artis) Stur; Calder et al., p. 184, Fig. 12A.
- 2006 Pecopteris plumosa (Artis) Brongniart; Wittry, p. 22, Figs. 1, 2; Fig. 3 (drawing after Dalinval 1960).
- T 2009 Senftenbergia plumosa (Artis) Stur; Cleal et al., Fig. 4 (copy of Artis’s original plate).
- 2016 Senftenbergia plumosa (Artis) Stur; DiMichele et al., p. 64, 67, Figs. 15a–15c.
- 2016 Senftenbergia plumosa (Artis) Stur; Opluštil et al., p. 169, 170, 171, 172, Figs. 14D, 15C.
- Excludenda:
- 1958 Pecopteris (Dactylotheca) plumosa (Artis) Kidston; Cross, p. 193, pl. 7, fig. 2 (difficult to judge from the figuration at natural size, but resembling Dicksonites plukenetii).
- 2017 cf. Senftenbergia plumosa (Artis) Stur; DiMichele et al., p. 54, fig. 30.3 (fragmentary and difficult to judge, but pinnules with different shape).
124 Description. Frond at least tripinnate. Antepenultimate rachis straight, flat, punctate, up to 6 mm wide. Pinna of penultimate order apparently lanceolate (incomplete), touching or slightly overlapping, at least 90 mm long. Penultimate rachis inserted at 60–80°, straight or slightly flexuous, rounded, smooth or finely striate longitudinally, ca. 1.5–2 mm wide. Last order pinnae alternate, closely spaced or slightly overlapping; linear-lanceolate, with distal pinnules gradually fusing into a blunt terminal; apical angle 35–45°. Dimensions: 15–40 mm long and 2–5 mm broad; length/breadth ratio = 7.5–8. Last order rachis inserted at 45–90°, straight, rounded, smooth or longitudinally striate, up to 0.7 mm wide. Pinnules alternate, obliquely inserted, united to the rachis by the entire basal width or even confluent, constituting a narrow wing along the rachis; pinnules subtriangular, with entire or slightly crenate margins and apices that vary from acute to broadly rounded; first basal catadromous pinnule slightly larger than the lateral ones; first anadromous pinnule smaller, with a more or less distinctly marked basal lobe. Dimensions: 4–9 mm long and 1–1.5 mm broad; length/breadth ratio = 4–6. Lamina thin, flat or slightly convex. Nervation well-marked. Midrib prominent, decurrent, slightly flexuous and extending into the apex; lateral veins straight, departing from midrib at a narrow angle; simple (in the smaller pinnules) or once forked (larger pinnules). Fertile pinnules similar in shape to the vegetative ones. Sporangia small, ovoid, placed upon lateral veins and occupying most of the space between midrib and pinnule margin.
125 Remarks. This variable but easily identifiable species was recorded by Bell (1944) as one of the most common and widespread plant fossils in the Cumberland Basin of Nova Scotia. Bell (1944) recorded the Canadian specimens as Pecopteris (Senftenbergia) plumosa forma crenata, highlighting the crenulate margin of pinnules, and contrasting it with Pecopteris (Senftenbergia) plumosa forma dentata, which has entire margins and more rounded apices; the latter would apparently characterize the upper Westphalian. However, he noted that both forms may be found in one and the same frond and that the distinction is one of degree, and thus of limited taxonomic use. Most authors regard the different forms of Senftenbergia plumosa as representing different parts of large fronds (e.g., Göppert 1836, Taf. XXVII; Stur 1885, Taf. XLV, fig. 1; Dalinval 1960, pl. VIII, fig. 1).
126 Comparisons.Senftenbergia pennaeformis has perpendicularly inserted, free pinnules of subrectangular shape, parallel margins, broadly rounded apex, and more sturdy aspect. Its venation is characterized by a thick and straight midrib giving off lateral veins that are once or twice forked at a wide angle and reach the pinnule margin nearly perpendicularly; these are embeded in a convex lamina. The shorter pinnules of Senftenbergia plumosa may be distinguished from those of Senftenbergia aspera by the thicker, convex lamina, more rounded apices, and more deeply imprinted veins in the latter.
127 Stratigraphic and geographic distribution.Senftenbergia plumosa is a very common, geographically widespread species; it is long-ranging, from upper Namurian (Yeadonian) through the entire Westphalian. The holotype originated from lower Duckmantian strata in the Yorkshire coalfield. It is most common in lower and middle Westphalian strata. Its stratigraphic usefulness is limited.
128 Occurrence in the Maritime Provinces, Canada. Cumberland Basin (Nova Scotia): Bell (1944): locality 858 (GSC 6536); locality 1031 (GSC 9444 + GSC 9445 ― fragmentary); locality 1050 (GSC 9210); locality 1053 (GSC 9947); locality 1086 (GSC 10055 ― fragmentary); locality 1087 (GSC 10123 ― fragmentary and poorly preserved; together with Dorycordaites palmaeformis + one piece without catalogue number ― together with Neuropteris obliqua, Alethopteris sp., Dorycordaites palmaeformis, Calamites sp., and Asterophyllites cf. longifolius); locality 1386 (GSC 9213); GSC 9209; locality 1481 (one piece without catalogue number ― fragmentary; together with rootlets); locality 1517 (one piece without catalogue number ― fragmentary); locality 2253 (one piece without catalogue number ― fragmentary and poorly preserved); locality 3110 (GSC 3076 ― together with Cordaites sp. and Samaropsis sp. + GSC 9200). Bell (1966): locality 1362 (GSC 15006). Calder et al. (2006). Fundy Geological Museum: FGM 999 GF 13.11/21.2 (together with Dorycordaites palmaeformis). Saint John (New Brunswick): locality 352 (128b ― labelled as Pecopteris serrulata); locality 788 (one piece without catalogue number); locality 2567 (two pieces, part and counterpart, without catalogue number ― both poorly preserved).
129 Occurrence in the United States. Illinois: Noé (1925), Andrews (1943), Janssen (1957), Langford (1958), Darrah (1969), Wittry (2006). Indiana: Canright (1959). Michigan: Arnold (1949). Missouri: Lesquereux (1879–80), White (1899), Basson (1968). Pennsylvania: Lesquereux (1879–80), Oleksyshyn (1982). Rhode Island: Lesquereux (1879–80). West Virginia: Jongmans (1937), Blake et al. (2002).
Order Coenopteridales Zimmermann 1930
Family Corynepteridaceae Cleal 1993
Genus Alloiopteris Potonié 1897
- 1893 Heteropteris Potonié, p. 44 (homonym — name introduced previously for extant plants).
- 1897 Alloiopteris Potonié, p. 138.
- 1956 Alloiopteris Potonié; Danzé, p. 94–96.
- 1983 Alloiopteris Potonié; Brousmiche, p. 84.
130 Type.Alloiopteris quercifolia (Göppert 1836) Potonié 1899.
131 Remarks.Alloiopteris refers to parallel-bordered last order pinnae with an approximately equal width throughout, and generally possessing narrowly confluent pinnules that are small and of virtually the same size and shape along the entire pinna length.
132 The name Alloiopteris is used for vegetative pinnules and pinnae that are similar in shape to fertile remains assignable to Corynepteris, or for specimens in which evidence for fertile status is uncertain; the distinction between Alloio pteris and Corynepteris is thus practical rather than definitive. Alloiopteris is widely distributed and relatively common, though, its occurrence is probably determined by environmental parameters. It ranges throughout the Pennsylvanian.
Alloiopteris almaensis Bell 1944
(Figs. 15a–b)
 Display large image of Figure 15
Display large image of Figure 15
- 1935 Alloiopteris (Corynepteris) coralloides forma compacta Gothan, p. 41, Taf. 41, fig. 2 (lower part).
- * v 1944 Alloiopteris almaensis n. sp. Bell, p. 75, pl. XXIV, fig. 2; pl. LXXI, fig. 2.
- ? 1944 Alloiopteris (Corynepteris) coralloides (Gutbier) Zeiller; Bell, p. 75, pl. XXII, fig. 4 (poorly preserved).
- 1956 Corynepteris coralloides var. compacta Gothan; Danzé, p. 312–313, pl. XLVII, figs. 4–6a.
133 Description and remarks. Bell (1944, p. 75) described a single pinna fragment of the penultimate order (holotype), as a new species, Alloiopteris almaensis. This species has been generally ignored by later authors.
134 Alloiopteris almaensis is characterized by pecopteroid pinnules joined at the base by a narrow strip of lamina associated with a slightly decurrent midrib; this imparts a slightly asymmetrical aspect to the pinnules. Basal catadromous pinnules are much more strongly developed (larger) than lateral ones, and thus stand out as individual elements. Lateral pinnules are variable in size along the pinna length, up to 5 mm long and 3 mm wide; they are weakly lobate, with angular lobes that appear bluntly pointed. Overall pinnule shape is subrectangular, with a rounded to bluntly pointed apex. The midrib is thin, decurrrent, flexuous and extending into the pinnule apex. Lateral veins are equally thin, once forked and widely spaced, with vein endings that seem to correspond to small projections on the pinnule margin.
135 Apart from the holotype, I include in Alloiopteris almaensis five additional fragmentary specimens from the type locality that have not been previously illustrated, as well as a specimen figured by Bell (1944, pl. XXII, fig. 4) as Alloiopteris(Corynepteris) coralloides, from the same area of New Brunswick. A more complete specimen from the Donald Reid Collection at Joggins, Nova Scotia, is also included and figured herein (Figs. 15a–b).
136 Bell (1944) compared Alloiopteris almaensis with Alloiopteris junghannii, a taxon introduced by Gothan (1913, p. 115–116, Taf. 25, figs. 4–5; Taf. 26, fig. 3) from the Upper Silesian Coal Basin. Two additional specimens of Alloiopteris junghannii from Langsettian strata of the same basin were figured by Stopa (1957, Tab. XX, figs. 1–2), and a further specimen was figured by Josten (1983, Taf. 38, figs. 2, 2a–b; Abb. 73) from the upper Namurian B (Marsdenian) of the Ruhr District, western Germany. Josten’s specimen shows relatively smaller (shorter) pinnules which are all of near-equal size. Gothan’s illustration at natural size does not show details of venation, but Beltz (1989, Taf. V, fig. 5; Taf. VI, figs. 1–1b) provided enlargements of the holotype. These show two kinds of pinnule. One type is similar to Alloiopteris sternbergii, displaying perpendicularly inserted, decurrent pinnules, united to the rachis by the entire base and narrowly confluent, of subquadrangular shape, with four short, pointed teeth on the distal margin; its venation consists of a single, decurrent vein that gives off a single lateral vein to each tooth. The other type of pinnule is subrectangular, more elongate, and has three to five pointed lobes with bifid, pointed teeth on the margin, the last feature apparently distinguishing it from Alloiopteris almaensis. However, larger specimens are required to establish the full range of morphological variation and whether one or two species are represented. Both Alloiopteris junghannii and Alloiopteris almaensis are known only from small fragments that are insufficient to show the intraspecific variation. In the absence of fertile remains, neither can be attributed to Corynepteris.
137 Beltz (1989) regarded Alloiopteris almaensis as synonymous with Alloiopteris pecopteroides, which was described by Gothan (1913, p. 110) from Upper Silesia. However, the fragmentary specimen figured by Gothan (1913, Taf. 25, figs. 2–2a; text-fig. 8) shows pinnules with a higher length/ breadth ratio as well as being of near-equal size throughout the pinna. The neotype designated by Beltz (1989, Taf. VI, figs. 2–2a), also shows more closely packed pinnules and a basal catadromous pinnule that is of similar size and shape to the lateral ones.
138 Alloiopteris almaensis most closely resembles a specimen figured as Alloiopteris coralloides forma compacta by Gothan (1935, Taf. 41, fig. 2 ― lower part) from Langsettian strata in the Ruhr District of Germany. Two other remains figured as Corynepteris coralloides var. compacta by Danzé (1956, pl. XLVII, figs. 4–6) from the Langsettian of Nord/Pas-de-Calais, northwestern France, also appear to represent the same taxon. All of these specimens show subrectangular, pecopteroid, weakly lobate pinnules with bluntly pointed apices. The nervation is also similar, with a thin, slightly decurrent, flexuous midrib extending into the tip of the pinnule. I include these European specimens in the synonmy with Alloiopteris almaensis.
139 Stratigraphic and geographic distribution.Alloiopteris almaensis is too rare and indistinctive to be useful stratigraphically. Bell’s (1944) specimens from New Brunswick, and those from the Donald Reid Collection from the Joggins section are from Langsettian strata and are the only known material from Canada. The European specimens recorded as Corynepteris coralloides forma compacta are also from the Langsettian.
140 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Donald Reid collection, Joggins, Nova Scotia (1999): DRC–997 64. Saint John (New Brunswick): Bell (1944): locality 1457 (GSC 9893 ― figured as Alloiopteris (Corynepteris) coralloides); locality 1461 (GSC 8569 ― holotype); locality 1461b (five pieces without catalogue number ― fragmentary).
Genus Corynepteris Baily 1860 emend. Galtier and Scott 1979
- 1860 Corynepteris Baily, p. 237–241.
- 1883 Saccopteris Stur, p. 64 [696] (based on Andrae’s Spheno pteris essinghii).
- 1883 Grand’Eurya Zeiller, p. 203 (based on Gutbier’s Spheno pteris coralloides).
- 1979 Corynepteris Baily; Galtier and Scott, p. 115.
- 1983 Corynepteris Baily; Brousmiche, p. 56–57.
141 Type.Corynepteris stellata Baily 1860.
142 Remarks.Corynepteris is characterized by perpendicularly inserted, parallel-sided last order pinnae of almost equal width throughout. Pinnules are small, attached to the rachis by the entire basal width, and generally confluent. Sporangia are large, ovate, sessile, and mostly with five sporangia in a sorus.
Corynepteris angustissima (Sternberg 1823) Němejc 1938
(Figs. 16a–c)
 Display large image of Figure 16
Display large image of Figure 16
- * 1823 Pecopteris angustissima Sternberg, p. 29, Taf. XXIII, figs. 1a–1b.
- * 1854 Asplenites Sternbergii Ettingshausen, p. 42, 73, Taf. 20, figs. 2–3; Taf. 20, fig. 4 (as Asplenites lindsaeoides in the plate explanation) (acc. to Němejc 1938).
- * 1884 Pecopteris Georgiana Lesquereux, p. 759–760, pl. XCVIII, figs. 6–6a (Lesquereux’ species has been rarely cited).
- 1900 Aloiopteris (sic) georgiana (Lesquereux) White, p. 776, 792, 793, 866, 883.
- 1923 Corynepteris (Zygopteris) Sternbergi, Kidston, p. 301–305, pl. LXXIV, figs. 1–3, 5, 5a–5d (spores); pl. LXXVIII, figs. 1–2.
- 1937 Alloiopteris Sternbergii, Jongmans, p. 405, pl. 25, fig. 66.
- 1938 Corynepteris angustissima, Němejc, p. 15, 41, pl. I, figs. 3–4; pl. III, figs. 8–10; text-figs. 4–5, 9b.
- v 1944 Alloiopteris (Corynepteris) sternbergi (Ettingshausen) Bertrand; Bell, p. 74, pl. XXIII, fig. 2 (herein Fig. 16a), fig. 3; pl. XXIV, fig. 1 (herein Figs. 16b–c).
- v 1944 Corynepteris sp.; Bell, p. 75–76, pl. XXV, figs. 1, 3.
- 1957 Corynepteris sternbergi (Ettingshausen) Zeiller; Janssen, p. 116, 117, Figs. 97, 99e (drawing).
- 1958 Corynepteris sternbergi (Ettingshausen) Zeiller; Langford, p. 256, Fig. 465.
- 1963 Alloiopteris sternbergii (Ettingshausen) Potonié; Cridland et al., p. 69, pl. 21, fig. 44 (magnificent specimen).
- v 1966 Alloiopteris (Corynepteris) sternbergi (Ettingshausen) Zeiller; Bell, p. 14, 16, 50, pl. VI, fig. 4; pl. VII, fig. 1; pl. XXIV, figs. 3–4.
- ? 1967 Corynepteris angustissima (Sternberg) Němejc; Tidwell, p. 36, pl. 5, figs. 2–3 (difficult to judge from the illustrations).
- ? 1969 Alloiopteris winslowii White; Darrah, p. 149, 215, pl. 22, fig. 1 (poorly figured).
- 1975 Alloiopteris sternbergi (Ettingshausen) Potonié; Jennings, p. 52–55, pl. 1, figs. 1–2 (adpression), figs. 3–9 (anatomy); text-fig. 1 (drawing).
- 1979 Alloiopteris angustissima (Sternberg) Potonié; Álvarez Ramis et al., p. 127–128, Taf. 1, figs. 1, 2; Abb. 1.
- 1979 Corynepteris sternbergii (Ettingshausen) Zeiller; Galtier and Scott, p. 89–97, pl. 1, fig. 6 (same as Kidston 1923, pl. LXXIV, fig. 5), fig. 7 (sporangia), fig. 8; pl. 2, figs. 1–7; pl. 3, figs. 1–11 (sporangia and spores); pl. 4, figs. 6, 8, fig. 7 (sporangia); text-figs. 2–5 (drawings).
- 1983 Corynepteris angustissima (Sternberg) Němejc; Brousmiche, p. 57–63 (including synonymy), pl. I, figs. 1–3 (same as Ettingshausen 1854, Taf. 20, figs. 2–4), figs. 4–5a, figs. 6–6a (same as Álvarez Ramis et al. 1979, Taf. 1, fig. 1), figs. 7–8a; pl. 2, figs. 1–2a, figs. 3–3a (same as Stur 1885, Taf. LII, fig. 3), figs. 4–6, figs. 7–7a (same as Gothan 1935, Taf. 43, fig. 2), figs. 8–8a (same as Stur 1885, Taf. LII, fig. 5); pl. 3; text-figs. 9A–C; text-fig. 10.
- 1984 Alloiopteris sternbergi (Ettingshausen) Potonié; Jennings, p. 307, pl. 3, fig. 5 (same as Jennings 1975, pl. 1, figs. 1–2).
- 1985 Alloiopteris Georgiana (Lesquereux) White; Gillespie and Crawford, p. 255, pl. III, fig. 8.
- ? 1996 Alloiopteris sternbergii (Ettingshausen) Potonié; Cross et al., p. 448, figs. 23–15.7, 23–15.8 (diffi-cult to judge from the illustrations).
- T 1997 Corynepteris angustissima (Sternberg) Němejc; Kvaček and Straková, p. 34, pl. 5, fig. 2 (photograph of holotype).
- 2002 Alloiopteris angustissima (Sternberg) Potonié; Blake et al., pl. XXXIV, fig. 4.
- 2011 Corynepteris sternbergii (Ettingshausen) Zeiller; Pšenička & Opluštil, p. 73–75, pl. VII, figs. 1–6.
- 2016 Corynepteris sternbergii (Ettingshausen) Zeiller; Opluštil et al., p. 171, Fig. 14B.
143 Description. Frond at least bipinnate. Penultimate rachis straight, rounded, punctate and with longitudinal striae, ca. 1.5–2 mm wide. Last order pinnae alternate, well spaced, parallel-sided and narrow, with rigid aspect. Dimensions: up to 35 mm long (incomplete) and 3 mm wide. Last order rachis inserted at right angle, straight, rounded, punctate and longitudinally striate, ca. 0.5 mm wide. Pinnules inserted perpendicularly, alternate, closely spaced, decurrent and united to the rachis by their entire basal width with a narrow confluence; subquadrate, markedly asymmetric, with four short, pointed teeth on the distal margin. Dimensions: 1–1.3 mm long and 0.9–1.1 mm wide; length/breadth ratio = 1.1–1.2. Lamina thick, convex. Nervation composed by a single, decurrent vein that gives off, at a wide angle, one lateral vein to each tooth.
144 Remarks. Bell (1944, pl. XXIII, figs. 2–3; pl. XXIV, fig. 1) figured as Alloiopteris (Corynepteris) sternbergi three fragmentary, but distinctive, specimens from Springhill, Nova Scotia. Bell (1966, pl. VI, fig. 4; pl. VII, fig. 1) figured another specimen from the Springhill area and one from the Sydney Basin (Bell 1966, pl. XXIV, figs. 3–4). Brousmiche (1983, p. 57) accepted Bell’s identification in her revision of the genus Corynepteris; Galtier and Scott (1979) did not mention Bell’s record.
145 I also include in Corynepteris angustissima the fragmentary specimen figured as Corynepteris sp. by Bell (1944, pl. XXV, figs. 1, 3). Although its preservation is poor, it seems characteristic of this species.
146 Comparisons.Alloiopteris erosa differs from Corynepteris angustissima in the general shape of its pinnules, which are subrectangular, obliquely inserted and strongly decurrent, with both lateral margins parallel to the rachis, as well as by the indentations of the upper margin. Its venation also differs in having a markedly decurrent midrib and forked lateral veins.
147 Stratigraphic and geographic distribution. Although never abundant, Corynepteris angustissima is a widely distributed, long-ranging species. It occurs from Chokierian (Namurian) to the uppermost Stephanian. The holotype is from uppermost Duckmantian strata of the Radnice Member, Kladno Basin, Bohemia, Czech Republic.
148 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Bell (1944): locality 881 (GSC 5970 ― together with Calamites suckowii + GSC 5971 ― both specimens figured as Corynepteris sp.); locality 1039 (GSC 9040 + GSC 9364 ― figured as Alloiopteris (Corynepteris) sternbergi + four pieces without catalogue number; together with Alethopteris urophylla, Zeilleria avoldensis, Renaultia schatzlarensis and Calamites sp.); locality 1052 (GSC 9426 ― figured as Alloiopteris (Corynepteris) sternbergi; together with rootlets + one specimen in the counterpart of GSC 8619 ― together with Renaultia footneri and Cordaites sp.); locality 1075 (GSC 9624 ― together with Stigmaria ficoides); locality 1982 (one fragmentary specimen without catalogue number) + GSC 9891 (figured as Alloiopteris (Corynepteris) sternbergi). Bell (1966): locality 1052 (GSC 15053 ― figured as Alloiopteris (Corynepteris) sternbergi; together with rootlets). Donald Reid collection, Joggins, Nova Scotia (1999): DRC 99 150 (together with Cordaites principalis). Sydney Basin (Nova Scotia): Bell (1966): locality 1338 (GSC 15044 ― figured as Alloiopteris (Corynepteris) sternbergi).
149 Occurrence in the United States. Georgia: Lesquereux (1884), Gillespie and Crawford (1985). Illinois: Janssen (1957), Langford (1958), Darrah (1969), Jennings (1975), Jennings (1984). Kansas: Cridland et al. (1963). Pennsylvania: White (1900). West Virginia: Jongmans (1937), Blake et al. (2002).
Order uncertain
Genus Discopteris Stur 1883
- 1883 Discopteris Stur, p. 61–64 [693–696].
- 1956 Discopteris Stur; Danzé, p. 58–60, 264–267.
- 1978 Discopteris Stur; Pfefferkorn, p. 170–172.
- 1983 Discopteris Stur; Brousmiche, p. 279–280.
150 Type.Discopteris karwinensis Stur 1883.
151 Remarks. Fertile pinnules characterized by annulate sporangia; sori rounded to oval, placed at the end of lateral veins or in the middle of lobes. Vegetative pinnules sphenopteroid, more or less lobate, joined to the rachis by the entire basal width or part of it. First basal catadromous element in ultimate, penultimate and antepenultimate order pinnae is a deeply segmented aphlebia.
cf. Discopteris vuellersii Stur 1885
(Fig. 17)
 Display large image of Figure 17
Display large image of Figure 17
- 1883 Discopteris Vüllersi Stur, p. 63, 64 [695, 696] (name only).
- * 1885 Discopteris Vüllersi Stur, p. 156, Taf. XXXIII, fig. 3.
- T 1978 Discopteris vuellersi Stur; Pfefferkorn, p. 183–188, Taf. 15, fig. 10 (Stur’s holotype), figs. 11–13 (details of holotype); Abb. 8–9.
- T 1983 Discopteris vuellersii Stur; Brousmiche, p. 284–287, pl. 77, figs. 1–1e (details of holotype), figs. 2–2a.
- v 1995 Discopteris vuellersii Stur; Álvarez-Vázquez, p. 196–198, lám. 66, figs. 1–3a.
152 Remarks. I tentatively include in Discopteris vuellersii a fragmentary but otherwise well-preserved specimen from Springhill, Nova Scotia that has not been previously illustrated. Discopteris vuellersii is a Westphalian species that is easily recognizable by its asymmetrical pinnules with bifid, deeply incised, acute lobes.
153 Stratigraphic and geographic distribution. Although rarely cited, Discopteris vuellersii is known from Langsettian to lower Asturian strata. The holotype is from Duckmantian strata of the Intra-Sudetic Basin of Lower Silesia.
154 Occurrence in the Maritime Provinces, Canada. Cumberland Basin (Nova Scotia): Bell (1944): locality 1031 (GSC9441 ― together with fragmentary remains of Alethopteris sp., Zeilleria sp. and Calamites sp.).
Genus Sphenopteris Brongniart 1822 sensu lato
- 1822 Sphenopteris Brongniart, p. 233.
155 Remarks.Sphenopteris is a problematic genus. Although the type, Sphenopteris elegans, is a Mississippian seed plant, the name is used also in a broader sense for ferns for which the reproductive structures are unknown and the systematic position is uncertain. This is the case for the species of Spheno pteris described below.
Sphenopteris cuneoliformis Bell 1944
(Figs. 18a–b)
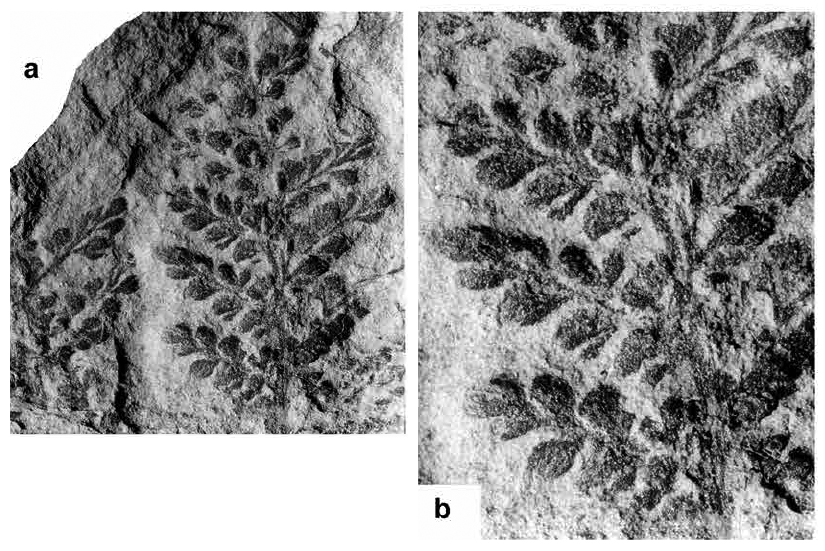 Display large image of Figure 18
Display large image of Figure 18
- * 1944 Sphenopteris cuneoliformis n. sp. Bell, p. 64, pl. VII, figs. 2–3 (herein Figs. 18 a–b).
156 Description. Bipinnate fragment preserved as an adpression on silty mudrock that is too coarse-grained for the outline of pinnules to be fully preserved. Last order pinnae lanceolate, closely spaced or only slightly touching laterally. Last order pinnae are exceedingly small, up to 8 mm long and 3.5 mm wide, with up to four pairs of lateral pinnules and a simple, distinct apical segment. Last order rachis inserted at ca. 45°, straight, rounded, thin, ca. 0.2 mm wide. Pinnules inserted by a short footstalk, ca. 1.1–1.5 mm long, almost isodiametric, entire and subcuneate, with a rounded apex in the smaller pinnules, and split into basal and terminal cuneate lobes in the larger ones. Lamina relatively thick, vaulted. No venation visible.
157 Remarks. Bell (1944, pl. VII, figs. 2–3) introduced Sphenopteris cuneoliformis on the basis of a single, fragmentary specimen. Although adequately described, the illustration is poor, possibly leading later authors to overlook it. Bell (1944) did not provide any comparisons, probably regarding it as clearly distinct from other sphenopterids. Although a species based on fragmentary remains will always be of questionable value, Sphenopteris cuneoliformis is retained here provisionally.
158 Comparisons. Most closely resembling Sphenopteris cuneoliformis is a specimen with small, lanceolate last order pinnae and small, subcircular, stalked pinnules illustrated by Danzé (1956, pl. LXXXVI, figs. 1–1a) as Sphenopteris cf. parviloba. This specimen is from an unspecified locality in Nord, northwestern France. The “true” Sphenopteris parviloba was introduced in the same paper (Danzé 1956, p. 513–516, pl. LXXXIII, figs. 3–5a) and is from upper Bolsovian strata. It has small pinnules with a generally orbicular outline, united to the rachis along most of their basal width and divided into up to three short, rounded lobes separated by shallow sinuses. Sphenopteris parviloba and Sphenopteris cf. parviloba are clearly two different species.
159 Sphenopteris cuneoliformis also resembles Sphenopteris microcyclos, another species characterized by small, rounded, more or less isodiametric pinnules. Poorly illustrated by its author (Gothan 1941, Taf. 52, fig. 3), Sphenopteris microcyclos was redefined by Danzé (1956, pl. LXXVI, figs. 1–2), who illustrated better-preserved specimens that differ from Sphenopteris cuneoliformis only by having a denticulate border (as shown in Danze’s text-fig. 50) (note that denticulations are not always evident in poorly preserved specimens). Gothan’s specimen is from the Duckmantian of the Upper Silesian Basin, whereas the specimens illustrated by Danzé are from middle to upper Bolsovian strata of Nord/Pas-de-Calais, France.
160 Sphenopteris cuneoliformis is also quite similar to specimens from Belgium figured as Hapalopteris microscopica by Stur (1885, Taf. XLIII, figs. 1–3). The latter species is characterized by small last order pinnae with very small pinnules, but the larger ones are trilobate.
161 Sphenopteris effusa possess similar pinnules, entire or three-lobed, isodiametric, oval to obovate in shape, with a rounded apex. However, these are stalked (as are those of Sphenopteris cf. parviloba), and pinnae and pinnules are of larger size. The holotype of Sphenopteris effusa (Lindley and Hutton 1937, pl. 204; Kidston 1923, pl. XVI, figs. 2–2a) is from lower Westphalian B (lower Duckmantian) strata of Felling Colliery, Durham, northern England. It shows a strongly developed rachis of the antepenultimate order that suggests a lower position in the frond than Sphenopteris cuneoliformis.
162 Stratigraphic and geographic distribution. Until now, Bell’s specimen is the only record of Sphenopteris cuneoliformis. It is from the Oxford area, Cumberland Basin, Nova Scotia.
163 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Bell (1944): locality 862 (GSC 5674).
Sphenopteris harttii Dawson 1862
(Fig. 19)
 Display large image of Figure 19
Display large image of Figure 19
- * 1862 Sphenopteris Harttii Dawson, p. 321, pl. XVI, figs. 48a–b (drawings).
- T 1868 Sphenopteris Harttii Dawson, p. 551, Fig. 192E (same as Dawson 1862, pl. XVI, fig. 48b).
- 1888 Sphenopteris Harttii Dawson, p. 73, Fig. 232E.
- T 1871 Sphenopteris Harttii Dawson, p. 52, pl. XVI, fig. 176 (same as Dawson 1862, pl. XVI, fig. 48b, and Dawson 1868, Fig. 192E), figs. 177–177a.
- p 1871 Hymenophyllites curtilobus Dawson; Dawson, p. 53, pl. XVI, fig. 178; non pl. XVI, figs. 179– 179a (indeterminable, fragmentary specimen with filiform lobes that Stopes 1914 indicated was lost).
- 1910 Sphenopteris Harttii Dawson; Matthews, p. 248 (name only).
164 Description and remarks. Here I describe previously unfigured, fragmentary remains of pinnae of penultimate order labelled as Sphenopteris harttii, a name introduced by Dawson (1862) for material from Saint John. Dawson’s original figures and description are poor, and no additional material of Sphenopteris harttii has been recorded subsequently. Only Schimper (1869, p. 409) and Kidston (1886, p. 227) briefly mentioned Sphenopteris harttii.
165 The specimens studied here (two pieces, part and counterpart) are up to 29 mm long and 25 mm wide, show pecopteroid, decurrent pinnules joined at the base by a strip of lamina; overall, they are subrectangular, with small, angular lobes; the first basal catadromous pinnule is smaller. Dimensions: 7–9 mm long and 2–3 mm wide. Midrib is decurrent, thin, although distinct in the thin lamina; it extends into the pinnule apex. Lateral veins are thin, widely spaced, departing from the midrib at a wide angle, with one lateral vein to each lobe.
166 Stopes (1914, p. 37, 105, 110) regarded Sphenopteris harttii as synonymous with Diplotmema subfurcatum (a pteridosperm, now included in Palmatopteris), whereas Bell (1944, p. 72) considered Sphenopteris harttii as synonymous with Zeilleria hymenophylloides. In my view, neither synonymy is correct.
167 As in the case of Sphenopteris cuneoliformis, and despite the problems associated with a species based on fragmentary remains, Sphenopteris harttii is retained here; as far as I know there is no other species comparable with this taxon.
168 Stratigraphic and geographic distribution. The material newly described here and Dawson’s (1862, 1868, 1871) material are from Saint John, New Brunswick; the material is thus within the range of upper Langsettian to middle Bolsovian age, but a more precise age has yet to be determined.
169 Occurrence in the Maritime Provinces, Canada. Saint John (New Brunswick): Dawson (1862, 1868, 1871). Geological Survey of Canada (material figured and described here): locality 117.
Sphenopteris cf. laciniosa Álvarez-Ramis et al. 1978
(Figs. 20a–b)
 Display large image of Figure 20
Display large image of Figure 20
- * 1978 Sphenopteris laciniosa Álvarez-Ramis et al., p.20, Taf. 9, fig. 6.
- T 1983 Sphenopteris laciniosa Álvarez-Ramis et al.; Brousmiche, p. 325–329, pl. 88, figs. 1–1a (holotype), figs. 2–7a; text-fig. 98.
170 Description. Frond at least tripinnate. Pinnae of penultimate order alternate, touching laterally or slightly overlapping. Dimensions: at least 40 mm long (incomplete) and 30 mm broad. Penultimate rachis inserted at 80–90°, straight, rounded, smooth, 0.5 mm wide. Last order pinnae alternate, closely spaced; subtriangular, with maximum width corresponding to the second pair of basal pinnules. Dimensions: 10–15 mm long and 5–6 mm wide; length/breadth ratio = 2–2.5. Last order rachis inserted at 70–80°, slightly flexuous, rounded, smooth, ca. 0.3 mm wide, narrowly winged due to confluent pinnule bases. Pinnules alternate, obliquely inserted, decurrent. Fully developed pinnules subquadrangular to subrectangular, obliquely inserted, with a slightly constricted base and composed of up to three pairs of bluntly pointed lateral lobes. Basal catadromous pinnule much shorter than the standard laterals (about half the length) and consisting of only three to four lobes. Dimensions: 3–5 mm long and 2.5–3 mm wide; length/breadth ratio = 1.2–1.6. Lamina thin, delicate. Venation well-marked, consisting of a ca. 0.1 mm width, slightly decurrent, nearly straight midrib that extends into the pinnule apex and gives off a lateral vein to each lobe.
171 Remarks. The unillustrated Canadian specimen studied here, stored at the Fundy Geological Museum, conforms to Sphenopteris laciniosa in the shape and dimensions of pinnules, their decurrent base, equidimensional growth, and the presence of bluntly pointed lobes. Although Brousmiche (1983) described a lesser development of the first basal catadromous pinnule of last order pinnae, these pinnules are still markedly shorter in the Canadian material, at half length, so the specimen cannot be assigned confidently and is designated as Sphenopteris cf. laciniosa.
172 Comparisons. In comparison with the Canadian material, Renaultia chaerophylloides has more sturdy pinnules with indented margins and more acute lobes. Fully developed pinnules of Renaultia schatzlarensis are of similar size to those of Sphenopteris cf. laciniosa and also show decurrent bases; but they are composed of five broader, more spreading, deeply incised, simple or bifid, obtusely pointed lobes.
173 Stratigraphic and geographic distribution.Sphenopteris laciniosa is rarely cited and never abundant. The holotype is from upper Bolsovian strata of Saarland, Germany. Brousmiche (1983) indicated that the species is characteristic of the Bolsovian.
174 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Fundy Geological Museum: FGM 999 GF 13.1/21.2 (together with Dorycordaites palmaeformis and rootlets).
Sphenopteris sp. A
(Figs. 21a–c)
 Display large image of Figure 21
Display large image of Figure 21
175 Description. Frond at least bipinnate. Penultimate rachis straight, flat, smooth, ca. 2.25 mm wide. Last order pinnae alternate, closely spaced; subtriangular, gradually tapering in the apical part to an elongate, subrounded, poorly individualized terminal. Dimensions: 35–120 mm long and 10–30 mm broad; length/breadth ratio = 3.5–4. Last order rachis inserted at right angle, rounded, smooth, ca. 0.5 mm wide. Pinnules alternate, subrectangular, elongate, obliquely inserted and broadly attached to the rachis (pecopteroid aspect); the larger ones show slightly lobed margins, rounded apex and a lobatopteroid venation. Dimensions: 5–12 mm long and 2–3 mm broad; length/breadth ratio = 2.5–4. Lamina thin, flat. Nervation composed by a slightly decurrent, very thin midrib, and lateral veins once or twice forked.
176 Remarks. The above description is based on a previously unfigured, unlabelled specimen from New Brunswick, illustrated herein as Figs. 21 a–c. Having been unable to establish its identity with any known species, I designate it provisionally Sphenopteris sp. A.
177 Comparisons. Compared with Sphenopteris sp. A, Senftenbergia plumosa has last order pinnae with higher length/ breadth ratio (7.5–8) and smaller subtriangular pinnules with entire or slightly crenate margins; the length/breadth ratio for Senftenbergia plumosa is higher (4–6) than for Sphenopteris sp. A. Lateral veins in Senftenbergia plumosa are straight, departing from midrib at a narrow angle; they are simple in smaller pinnules and bifurcate in the larger ones.
178 Occurrence in the Maritime Provinces, Canada. New Brunswick: locality 273.
Indeterminable fern fragment
- v 1944 Rhodea cf. sparsa, Bell, p. 57, pl. II, fig. 4; pl. XIII, fig. 3.
179 Remarks. The fragmentary and poorly preserved specimen figured as Rhodea cf. sparsa by Bell (1944, pl. II, fig. 4; pl. XIII, fig. 3) is here regarded as indeterminable. Re-examination of this specimen shows it to consist of two small pinnae with poorly preserved pinnules that certainly do not belong to Rhodea sparsa.
180 Occurrence in the Maritime Provinces, Canada. Cumber-land Basin (Nova Scotia): Bell (1944): locality 1564 (GSC 5938).
SUMMARY AND CONCLUSIONS
181 The record of Filicopsida from lower to middle Westphalian strata of Nova Scotia and New Brunswick, eastern Canada, derives from two sources: more or less random collecting during of field mapping by personnel of the Geological Survey of Canada; and material dumped as a result of coal mining. The record is thus incomplete, and most specimens are fragmentary. Early records by Dawson (1862, 1868) are particularly fragmentary. Nonetheless, the present revision, based mainly on the material recorded by Bell (1944, 1966) with additional specimens from the collections of the New Brunswick Museum at Saint John, the Fundy Geological Museum at Parrsboro, and the Joggins Fossil Institute (Donald Reid collection), has yielded a surprisingly large number of fern species. As with other Carboniferous plant groups, most species are the same as those recorded from western Europe, which was palaeogeographically close.
182 Bell’s identifications are generally sound, but some have needed revision, particularly the new species introduced by Bell (1944). Only two of his seven new species, Alloiopteris almaensis and Sphenopteris cuneoliformis, have been provisionally maintained after revision. Both species are based on fragmentary, inadequate specimens, and larger and better preserved material is needed for a better understanding. In addition, only one of the seven species introduced by Dawson, Sphenopteris harttii, is provisionally accepted herein. The other six species are compared with well-known European taxa. Although three of these species were first described by Dawson from Canada, it is necessary to wait the revision of Dawson’s material to confirm whether or not they are synonymous.
183 Table 1 lists the identifications accepted herein in comparison with those in Dawson (1862, 1868, 1871) and Bell (1944, 1966).
 Display large image of Table 1
Display large image of Table 1
John Utting is gratefully acknowledged for the initiative to promote a general revision of lower Westphalian floras in the Maritime Provinces of Canada, an update initiated by Robert H. Wagner in the late 1990s. I want to thank Roberto (Bob) for having infected me with his passion for work, which has allowed me to grow as a person, a palaeobotanist and a geologist. Jean Dougherty, Geological Survey of Canada, Ottawa, and Randall Miller, the New Brunswick Museum, Saint John, provided essential access and data regarding the fossil collections in their institutions. Donald Reid, Jog-gins Fossil Centre, kindly lent several specimens. The Fundy Geological Museum at Parrsboro lent the specimens from Spicer’s Cove. The helpful and well-focused suggestions of the two referees, Stanislav Opluštil and Josef Pšenička are gratefully acknowledged. The meticulous editing by Rob Fensome and all his recommendations to help improve the quality of this contribution are much appreciated.
APPENDIX
List of names of generic or lower rank of taxa cited herein, with authorships.
- Adiantites adiantoides (Lindley and Hutton 1834) Kidston 1923
- Alethopteris Sternberg 1825
- Alethopteris urophylla (Brongniart 1834) Göppert 1836
- Alloiopteris Potonié 1897
- Alloiopteris angustissima (Sternberg 1825) Potonié 1897
- Alloiopteris almaensis Bell 1944
- Alloiopteris coralloides (Gutbier 1835) Zeiller 1888
- Alloiopteris coralloides forma compacta Gothan 1935
- Alloiopteris erosa (Gutbier 1843) White 1899
- Alloiopteris georgiana (Lesquereux 1884) White 1900
- Alloiopteris junghannii Gothan 1913
- Alloiopteris pecopteroides Gothan 1913
- Alloiopteris quercifolia (Göppert 1836) Potonié 1899
- Alloiopteris sternbergii (Ettingshausen 1854) Potonié 1896
- Alloiopteris winslowii White 1899
- Annularia Sternberg 1821
- Annularia ramosa Weiss 1884
- Aspidites glockeri Göppert 1836
- Aspidites silesiacus Göppert 1836
- Asplenites lindsaeoides Ettingshausen 1854
- Asplenites sternbergii Ettingshausen 1854
- Asterophyllites Brongniart 1822
- Asterophyllites grandis (Sternberg 1825) Geinitz 1855
- Asterotheca Presl in Corda 1845
- Asterotheca arborescens (Schlotheim 1804 ex Brongniart 1828a) Stur 1877
- Asterotheca inferior Stockmans and Willière 1966
- Asterotheca praecursor Stockmans and Willière 1966
- Bertrandia Dalinval 1960
- Bertrandia avoldensis (Stur 1883) Dalinval 1960
- Boweria Kidston 1911
- Boweria minor Kidston 1923
- Boweria schatzlarensis Kidston 1911
- Boweria nowarudensis Frojdová et al. 2017b
- Calamites Suckow 1784
- Calamites carinatus Sternberg 1824
- Calamites suckowii Brongniart 1828a
- Callipteris Brongniart 1849
- Callipteris pilosa (Dawson 1868) Dawson 1871
- Calymmotheca Stur 1877
- Calymmotheca avoldensis Stur 1883
- Calymmotheca frenzli Stur 1883
- Calymmatotheca Zeiller 1883
- Cordaites Unger 1850
- Cordaites principalis (Germar 1848) Geinitz 1855
- Corynepteris Baily 1860
- Corynepteris angustissima (Sternberg 1823) Němejc 1938
- Corynepteris coralloides var. compacta Danzé 1956
- Corynepteris stellata Baily 1860
- Corynepteris sternbergii (Ettingshausen 1854) Zeiller 1899
- Corynepteris winslovii White 1899
- Crossotheca Zeiller 1883
- Crossotheca boulayi Zeiller 1888
- Crossotheca bourozi Danzé 1956
- Crossotheca compacta Bell 1938
- Crossotheca crepinii Zeiller 1883
- Crossotheca kidstonii Hemingway ex Crookall 1930
- Crossotheca schatzlarensis (Stur 1883) Kidston 1888
- Cyperites Lindley and Hutton 1832
- Dactylotheca plumosa (Artis 1825) Kidston 1886
- Dactylotheca plumosa forma dentata Bell 1938
- Dicksonites plukenetii (Schlotheim ex Sternberg 1825) Sterzel 1881
- Diplotmema furcatum (Brongniart 1829) Stur 1877
- Diplotmema sanctifelicis Stur 1885
- Discopteris Stur 1883
- Discopteris karwinensis Stur 1883
- Discopteris vuellersii Stur 1883
- Dorycordaites palmaeformis (Göppert 1852) Grand’Eury 1877
- Eusphenopteris pulchrior (Gothan and Jongmans 1915) Amerom 1975
- Eusphenopteris sauveurii (Crépin 1880) Simson-Scharold 1934
- Filicites plumosus Artis 1825
- Germera Brousmiche 1983
- Germera brousmicheae sp. nov.
- Germera mendescorreae (Teixeira 1939) Brousmiche 1983
- Grand’Eurya Zeiller 1883
- Hapalopteris Stur 1883
- Hapalopteris amoena Stur 1885
- Hapalopteris bella Stur 1885
- Hapalopteris aschenborni Stur 1885
- Hapalopteris microscopica Crépin ex Stur 1885
- Hapalopteris schatzlarensis Stur 1885
- Hapalopteris typica Stur 1883
- Heteropteris Potonié 1893
- Hymenophyllites curtilobus Dawson 1862
- Hymenotheca dathei Potonié 1890
- Karinopteris obtusifolia (Gothan 1941) Álvarez-Vázquez 1995
- Kidstoniopteris Frojdová et al. 2017b
- Kidstoniopteris minor (Kidston 1923) Frojdová et al. 2017b
- Laveineopteris polymorpha (Dawson 1862) Wagner 2008
- Lepidostrobophyllum Hirmer 1927
- Lyginopteris hoeninghausii (Brongniart 1830) Gothan 1931
- Neuropteris serrulata Dawson 1862
- Oligocarpia Göppert 1841
- Oligocarpia brongniartii Stur 1883
- Oligocarpia gutbieri Göppert 1841
- Oligocarpia mixta (Schimper 1869) Abbott 1954
- Oligocarpia splendens (Dawson 1871) Stopes 1914
- Oligocarpia stipulataeformis Stur 1885
- Palmatopteris Potonié 1892
- Palmatopteris alata (Brongniart 1830) Potonié 1892
- Palmatopteris furcata (Brongniart 1829) Potonié 1892
- Paripteris Gothan 1941
- Pecopteris Brongniart 1828a
- Pecopteris alloiophylla Doubinger and Germer 1971
- Pecopteris angustifida Ettingshausen 1854
- Pecopteris angustissima Sternberg 1823
- Pecopteris avoldensis (Stur 1883) Corsin 1932
- Pecopteris brachyphylla Doubinger and Germer 1971
- Pecopteris chaerophylloides Brongniart 1836
- Pecopteris clarkii Lesquereux 1880
- Pecopteris delicatula Brongniart 1836
- Pecopteris densifolia Dawson 1871
- Pecopteris dentata Brongniart 1836
- Pecopteris engyphylla Doubinger and Germer 1971
- Pecopteris georgiana Lesquereux 1884
- Pecopteris glockeriana Presl in Sternberg 1838
- Pecopteris micromiltonii Bertrand ex Corsin 1951
- Pecopteris miltonii (Artis 1825) Brongniart 1828a
- Pecopteris mucronata Presl in Sternberg 1838
- Pecopteris pachyneura Doubinger and Germer 1971
- Pecopteris pennaeformis Brongniart 1828a
- Pecopteris pilosa (Dawson 1868) Bell 1944
- Pecopteris plumosa (Artis 1825) Brongniart 1832
- Pecopteris plumosa var. dentata Sterzel 1901
- Pecopteris plumosa forma crenata Bell 1944
- Pecopteris preciosa Hartt in Dawson 1868
- Pecopteris pseudovestita White 1899
- Pecopteris pycnoneura Doubinger and Germer 1971
- Pecopteris serra Lindley and Hutton 1834
- Pecopteris serrulata (Dawson 1862) Dawson 1868
- Pecopteris silesiaca Göppert 1845
- Pecopteris tenuinervosa Corsin 1951
- Renaultia Zeiller 1883
- Renaultia chaerophylloides (Brongniart 1836) Zeiller 1883
- Renaultia crepinii (Stur 1885) Zeiller 1899
- Renaultia footneri (Marrat 1872) Kidston 1893
- Renaultia gracilis (Brongniart 1828a) Zeiller 1883
- Renaultia hemingwayi Kidston 1923
- Renaultia hydei Bell 1944
- Renaultia rotundifolia (Andrae 1869) Zeiller 1899
- Renaultia schatzlarensis (Stur 1885) Kidston 1890
- Rhodea Presl in Sternberg 1838
- Rhodea laqueata Bell 1944
- Rhodea sparsa Kidston 1923
- Rhodea stachei Stur 1877
- Rhodeopteridium Zimmermann 1959
- Saccopteris Stur 1883
- Samaropsis Göppert 1864
- Senftenbergia Corda 1845
- Senftenbergia aspera (Brongniart 1828a) Stur 1877
- Senftenbergia pennaeformis (Brongniart 1828a) Stur 1877
- Senftenbergia plumosa (Artis 1825) Stur 1877
- Sorotheca Stur 1883
- Sphenophyllum cuneifolium (Sternberg 1823) Zeiller 1879
- Sphenopteris (Brongniart 1822) Brongniart 1828a
- Sphenopteris aculeata Bell 1838
- Sphenopteris alata Brongniart 1830
- Sphenopteris amoenaeformis Kidston 1923
- Sphenopteris bella (Stur 1885) Zeiller 1899
- Sphenopteris boehnischii (Stur 1883) Šusta 1928
- Sphenopteris boulayi Zeiller 1886
- Sphenopteris bronnii Gutbier 1835
- Sphenopteris cantiana Kidston 1923
- Sphenopteris caudata Lindley and Hutton 1832
- Sphenopteris coralloides Gutbier 1835
- Sphenopteris crenata Lindley and Hutton 1834
- Sphenopteris crepinii Zeiller 1883
- Sphenopteris cuneoliformis Bell 1944
- Sphenopteris delicatula Sternberg 1825
- Sphenopteris deltiformis Kidston 1914
- Sphenopteris digitata Doubinger 1959
- Sphenopteris dixonii Kidston 1917
- Sphenopteris effusa Kidston 1886
- Sphenopteris essinghii Andrae 1866
- Sphenopteris fletcheri Bell 1944
- Sphenopteris footneri Marrat in Higgins 1872
- Sphenopteris gracilis (Brongniart 1828a) Zeiller 1888
- Sphenopteris harttii Dawson 1862
- Sphenopteris hoeninghausii Brongniart 1828a
- Sphenopteris hymenophylloides (Kidston 1923) Gothan 1941
- Sphenopteris laciniosa Álvarez Ramis et al. 1978
- Sphenopteris laurenti Andrae 1865
- Sphenopteris marginata Dawson 1862
- Sphenopteris mendescorreae Teixeira 1939
- Sphenopteris microcyclos Gothan 1941
- Sphenopteris mixta Schimper 1869
- Sphenopteris moyseyi Kidston 1923
- Sphenopteris parviloba Danzé 1956
- Sphenopteris philipensis Bell 1944
- Sphenopteris pilosa Dawson 1868
- Sphenopteris quadriloba Kidston 1923
- Sphenopteris rotundifolia Andrae 1869
- Sphenopteris rutaefolia Gutbier 1835
- Sphenopteris sampaiana Teixeira 1939
- Sphenopteris sanctifelicis (Stur 1885) Rydzewski 1913
- Sphenopteris schatzlarensis (Stur 1885) Zeiller 1899
- Sphenopteris schilleri Gothan 1913
- Sphenopteris schwerinii (Stur 1885) Zeiller 1899
- Sphenopteris souichii Zeiller 1888
- Sphenopteris splendens Dawson 1871
- Sphenopteris stipulataeformis (Stur 1885) Gothan 1913
- Sphenopteris philipensis Bell 1944
- Sphenopteris pilosa Dawson 1868
- Sphyropteris Stur 1883
- Sphyropteris boehnischii Stur 1883
- Sphyropteris crepinii Stur 1883
- Sphyropteris frankiana Gothan 1912
- Sphyropteris obliqua (Marrat 1872) Kidston 1889
- Sphyropteris tomentosa Stur 1883
- Stigmaria ficoides (Sternberg 1820) Brongniart 1822
- Sturia Němejc 1934
- Sturia amoena (Stur 1885) Němejc 1934
- Urnatopteris herbacea (Boulay 1876) Carpentier 1919
- Zeilleria Kidston 1884
- Zeilleria avoldensis (Stur 1883) Kidston 1884
- Zeilleria delicatula Kidston 1884
- Zeilleria frenzlii (Stur 1883) Kidston 1884
- Zeilleria hymenophylloides Kidston 1924
- Zeilleria lignyi Stockmans and Willière 1958
- Zeilleria pilosa (Dawson 1868) Wagner 2001b
- Zeilleria schaumburglippeana (Stur 1885) Zeiller 1907
- Zeilleria stellata Arnold 1937