Late Prehistoric Human Impact on Bass Pond, Port au Choix
Trevor BellMemorial University
Joyce B. Macpherson
Department of Geography, Memorial University
M. A.P. Renouf
Archaeology Unit of Memorial University
INTRODUCTION
1 CALIBRATED RADIOCARBON DATES place the Groswater and Dorset Palaeoeskimo occupations of Port au Choix at 2950-1820 cal BP1 and 1990-1180 cal BP, respectively. Faunal studies (Kennett 1990; Murray 1992; Hodgetts et al. 2003) have demonstrated that populations of both cultures engaged in harp seal hunting during the late winter and early spring, and Hodgetts (this volume) demonstrates that Dorset also exploited harp seal during the early winter. Excavation of Groswater and Dorset sites at Port au Choix (Renouf 1993, 1994, 1999) reveals that Dorset occupation was more intensive than Groswater, and was more narrowly focused on the harp seal.
2 In this paper we demonstrate that at 2200-1800 cal BP there were marked changes in the local environment observable in the fossil pollen and spores, algal remains and charcoal in pond sediments from Bass Pond, adjacent to Palaeoeskimo sites in Port au Choix (Figure 1). We compare these to data from Stove Pond, which represents a prehistoric baseline record from an interior, non-inhabited location. The changes observable in the Bass Pond environmental record are coincident with nearby Palaeoeskimo occupations, and we argue that Groswater and Dorset activities had clear and recognizable impacts on their local environment through (i) alteration of the natural vegetation by wood-cutting and burning; (ii) disturbance and erosion of soil; (iii) disturbance and trampling of wetlands; and (iv) temporary increases in lake nutrients (eutrophication) and salinity.
Figure 1. Location map for Bass and Stove ponds and place names used in the text.
CULTURAL CONTEXT
3 Bass Pond is adjacent to two important Palaeoeskimo sites and within a half-kilometre of a third (Figure 2). Phillip’s Garden East is a 1500-square-metre Groswater Palaeoeskimo harp seal hunting site, as shown by faunal material and hunting equipment. There are two migratory periods when harp seal are available in this area: early winter and late winter-early spring (Sergeant 1991). LeBlanc (1996) argues that the late winter-early spring migration was the more predictable, and therefore the more important of the two hunting seasons.
4 There are two temporal and spatial areas of occupation at Phillip’s Garden East which together span 2950-2130 cal BP. The earlier occupation area has three dates at 2950-2490 cal BP and a fourth date at 2650-2150 cal BP (Table 1). The later occupation area has seven dates at 2720-2130 cal BP and includes remains of a tent structure, defined by a low perimeter berm surrounding a level area, and two cache pits with stone covers (Renouf 1994). Both areas are littered with fire-reddened and -cracked rock, reflecting the primary mode of heating by hot rocks, rather than the oil lamps characteristic of later Dorset people. The faunal assemblage is predominantly seal, although it included a range of small game and bird in small proportions (Kennett 1990; Renouf 1994). The tool assemblage from both areas is relatively narrow, focused on tools for hunting and processing activities, and without much evidence of tool making (Kennett 1990; Renouf 1994; LeBlanc 1996). There are eighteen tools that were originally interpreted as axes (Renouf 1994). Upon closer inspection, although four of these are clearly axes, and as such reflect wood-cutting and wood-working activities, the remaining fourteen are smaller and could have been more multi-functional, analogous to the Inuit curved knife or ulu (Figure 3). Amongst the many functions of the ulu was skin scraping (Rankin and Labreche 1991), which would have been a logical component of the harp seal hunting and processing activities carried out at the site. The small size of the site, the insubstantial nature of the dwelling, and the relative narrowness of the tool assemblage suggest that Phillip’s Garden East was regularly occupied for a few weeks or months during the late winter, for the primary purpose of harp seal hunting. If our interpretation of the tools in Figure 3 is correct, it suggests that sealskin processing activities might have gone on as well.
Figure 2. Oblique aerial photograph looking southeast across Point Riche Peninsula to Port au Choix in the background. Phillip’s Garden, in the foreground, is built on a series of raised beaches that extend up to the base of Crow Head and form a broad arc behind the site. Bass Pond is located 500 m to the east of Phillip’s Garden.

Display large image of Figure 2
5 Phillip’s Garden East is connected to another Groswater site less than one-half kilometre away, Phillip’s Garden West. This site is situated on a small (500 square metre) high terrace with a panoramic ocean view. A lightweight tent structure is inferred from a ring of small post-holes for narrow poles that would not have borne much weight, and refuse was thrown down the hillside. Faunal material from this hillside midden reflects the late winter-early spring harp seal hunt (Wells this volume). Eight dates place the occupation at 2680-1820 cal BP, thus overlapping with the younger occupation area at Phillip’s Garden East and persisting beyond that occupation as presently dated; there is the logical possibility that further excavation at Phillip’s Garden East would extend that site’s temporal range. Renouf (2005) divided Phillip’s Garden West into two spatially and temporally distinct areas, the hillside midden and the upper terrace. Three dates from the midden area place it at 2680-2160 cal BP with a fourth date at 2000-1820 cal BP. Three dates from the upper terrace place it at 2340-1950 cal BP and a fourth date is at 2680-2210 cal BP. Wells (2002) argues that Phillip’s Garden East and West were connected during their period of chronological overlap (2680-2150 cal BP) through coordinated activities relating to the hunting and processing of seal.
Table 1. Radiocarbon dates and calibrated calender ages (1σ) for samples collected from archeological sites at Philip's Garden East, Philip's Garden West and Philip's Garden. and sediment cores from Stove and Bass ponds. Radiocarbon dates were calibrated using Calib 4.4 (Stuiver and Reimer 1993)
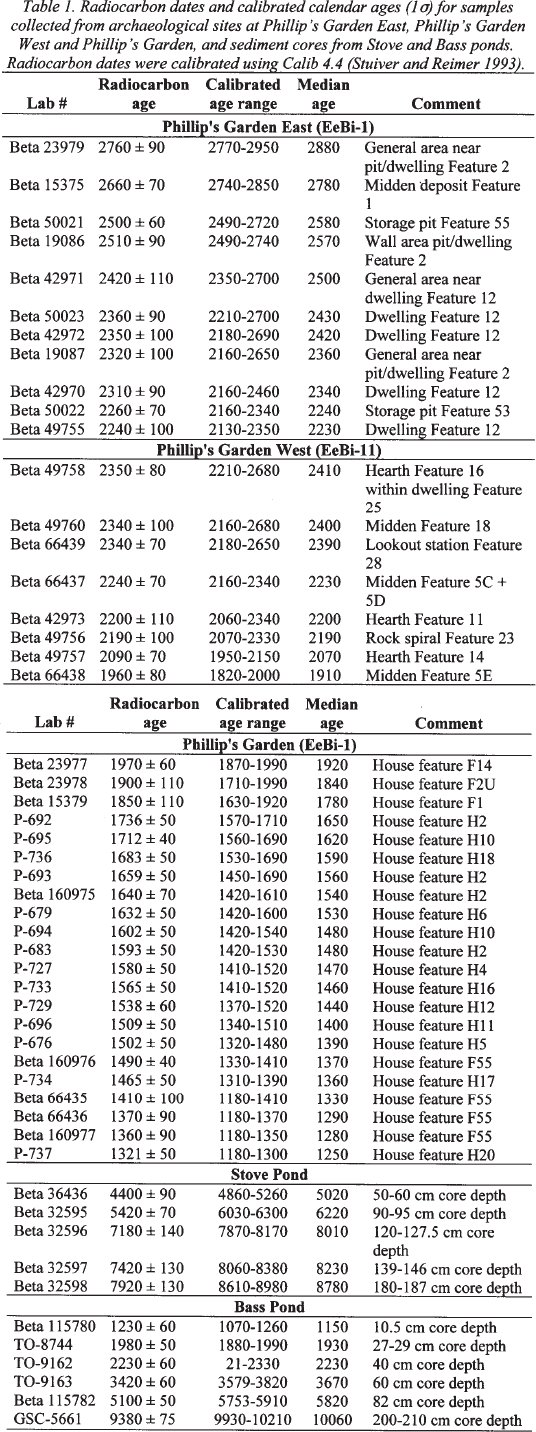
Display large image of Table 1

Display large image of Figure 3
6 In contrast to these Groswater occupations, the succeeding Dorset Palaeoeskimo occupation of Phillip’s Garden (1990-1180 cal BP) was much more intensive. The remains of 68 dwellings are identified at this large site and there are likely many more obscured by post-abandonment midden in-fill and the encroaching tuckamore (stunted spruce scrub forest). Midden and dwelling assemblages are very rich in both faunal and artefactual material, reflecting the large amount of garbage accumulation which in turn reflects the intensity of human occupation at the site. While the taxonomic diversity of the faunal remains is limited, with an almost exclusive focus on harp seals (Harp 1976; Renouf 1999; Hodgetts et al. 2003), the artifactual material reflects a wide range of hunting, domestic and spiritual activities (Renouf 2004). The tool assemblage includes harp seal hunting equipment (bone harpoons and stone endblades), butchering tools (chert microblades and bifacial knives) and seal skin processing equipment (ground slate scrapers, chert scrapers, bone awls).
7 On the basis of radiocarbon-dated dwellings, Harp (1976) and Erwin (1995) characterize site evolution as an initial small population, followed by an increase in population and a subsequent decline. We date these three phases of site development at 1950-1550 cal BP, 1550-1350 cal BP, and 1350-1180 cal BP (Figure 4). Throughout all phases, features suggest that the Phillip’s Garden dwellings were considerably larger and more substantially constructed than the Groswater dwelling structures (Renouf 2004).
8 Did this lengthy Palaeoeskimo occupation of the Bass Pond area (2950-1180 cal BP) change the local vegetation in any way? Human impact might be seen in the changing vegetation and pond water conditions inferred from proxy data such as fossil pollen and spores, algal remains and charcoal from Bass Pond sediments (Figure 5).
POLLEN ANALYSIS AND HUMAN IMPACTS IN THE BOREAL FOREST
9 Vegetation changes identified through the analysis of pollen records from the bo-real forest of Canada have largely been ascribed to natural factors, such as climate. In boreal Europe, where human influence on natural vegetation has been far more profound, pollen analysts have been more ready to examine its effects (e.g., Behre 1986), for example by identifying the impacts of Neolithic agriculture on the forest.
10 Throughout much of western Europe, the transition from nomadic hunter-gathering or semi-nomadic herding to settled agriculture occurred millennia ago. However, in northern Scandinavia the Saami people still maintain to some extent a culture based on semi-nomadic reindeer herding. Examination of modern, or historically recent pollen assemblages from this area has provided comparative information that has been used to identify small-scale prehistoric impact upon the forest (Hicks 1993; Aronsson 1994). A distinctive suite of herb and low shrub pollen taxa characterized the recently disturbed sites, with accompanying reductions in the proportions of the dominant forest trees. A similar pollen signature was found in a pollen profile from an archaeological site in northern Sweden, where Aronsson (1994) was able to identify two phases of semi-nomadic occupation. These were marked by minor decreases in the proportions of tree pollen and small increases in the pollen of herbs, most notably Poaceae (grasses), with smaller amounts of Rumex (dock), Artemisia (wormwood), Ranunculaceae (buttercup), and Rosaceae.
Figure 4. Dated dwellings at Phillip’s Garden. H# refers to a dwelling excavated by Harp (1976) and F# refers to a dwelling excavated by Renouf (2002). The vertical bar represents the 1-sigma probability range for the calibrated radiocarbon date. The small circle within each bar represents the median probability calendar age.
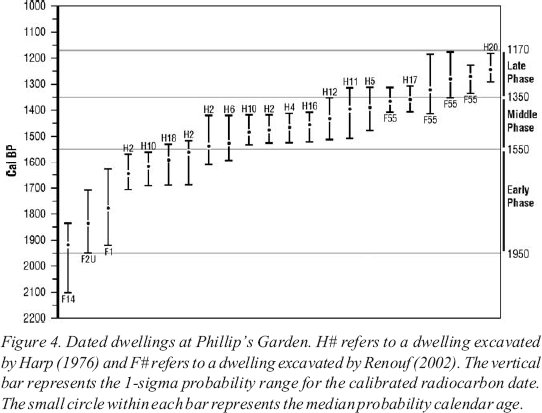
Display large image of Figure 4
11 Hicks (1993) identified more profound changes in pollen from peat at a site in northern Finland, occupied in winter in the fifteenth and seventeenth centuries. The changes included a marked decrease in the pollen of pine, the dominant forest tree, and increases in Ericales (heaths), Poaceae, other herbs, and charcoal. The period of disturbance was followed by a period with high values for birch pollen before the regional pine forest was re-established. Hicks (1993) interprets this sequence of changes as indicating first an increase in heaths as the forest became more open, then an increase in herbs, especially grasses, as disturbance became more severe. Increases of charcoal and of the pollen of Chamerion (Epilobium: fireweed) suggested small local fires. Abandonment of the site led to forest regeneration, first by birch, both tree and shrub, which was gradually replaced by the regional pine.
Figure 5. A sample of representative pollen grains and spores identified from Bass Pond sediment core.
Display large image of Figure 5
12 Many boreal trees are wind-pollinated and therefore prolific producers of easily dispersed pollen, whereas the herbs and shrubs growing on the forest floor and on disturbed or open ground usually produce relatively small amounts of pollen that is less readily dispersed. Some, such as the Ericales, are insect-pollinated. Therefore, most pollen assemblages are dominated by pollen from wind-dispersed plants and often contain a considerable proportion of pollen derived from far distant sources. Nevertheless, the Scandinavian workers, using pollen assemblages derived from moss or peat samples, found that disturbance was expressed in the pollen only close to a disturbed (archaeological) site. Therefore, only peat samples collected close to a site would be expected to record the signal of the disturbance. In contrast, samples from lake sediment, as in this paper, integrate the pollen rain from a wider area, and can provide a broader if more diffuse regional signal.
STOVE POND AS A CONTROL SITE
13 To evaluate the effects of prehistoric human disturbance on vegetation it is necessary to establish baseline data from an undisturbed site. In the Port au Choix area this site is the 4-hectare Stove Pond which lies on the coastal plain 11 kilometres ESE of Port au Choix at an elevation of about 55 metres (Figure 1; Macpherson 1997). It is located on an inlier of Early Cambrian sandstone (Macpherson 1997).
14 Stove Pond lies between a raised bog to the west dominated by the mosses Sphagnum and Cladonia and a moraine ridge to the east rising to about 70 metres above sea level and supporting Abies balsamea-dominated forest (Bouchard et al. 1978; Grant 1994). Stream banks nearby support thickets dominated by Alnus rugosa. A sediment core was obtained from the lake in July 1987 in a water depth of1.43 metres. The top 105 centimetres of the 253-centimetre core was composed of highly organic mud (>80% loss at 500EC), the base of the core consisted of clay (Macpherson 1997).
Pollen diagram: explanation
15 A summary pollen diagram from the organic sediment from the Stove Pond core is given in Figure 6. The principal y-axis shows the age of the sediment in calibrated (calendar) years before present (cal BP), derived from five radiocarbon dates on bulk organic sediment (Table 1). Only the period covering prehistoric human occupation — the last 7000 years — up to the present is reproduced here. The secondary y-axis shows sediment depth; variations in the scale indicate a reduction in the rate of sediment accumulation since about 5000 cal BP. Percentages of individual taxa or groups of taxa are shown on the x-axis, with values of less than 10% shown with an exaggerated scale. Percentages of pollen grains are shown in black, with the types grouped according to the nature of the taxa. It must be borne in mind, however, that some tree species are dwarfed in tuckamore (conifer scrub) or on bogs; the nature of the pollen produced by these small plants is unaltered but the quantity is much reduced. The final curve shows the concentration of pollen per millilitre of sediment, a measure varying with the pollen productivity of the contributing vegetation and with the rate at which the sediment accumulated.
16 Percentages for spores (mostly ferns and fern allies) are shown in a striped pattern; these percentages are calculated “outside the sum” (the sum for these taxa is the pollen sum plus the total spore sum). Concentrations of Pediastrum (an aquatic alga and indicator of the nutrient status and light transmissivity of the lake water) and charcoal fragments (evidence of fire) are shown as histograms. Microscopic charcoal fragments can be carried for long distances by the wind and do not necessarily indicate fires in the lake’s catchment (Cwynar 1978). Indeed, forest fires are very infrequent on the western coastal plain of the Great Northern Peninsula of Newfoundland (Meades and Moores 1989).
Figure 6. Summary pollen percentage diagrams for the last 7000 cal BP from Stove and Bass ponds.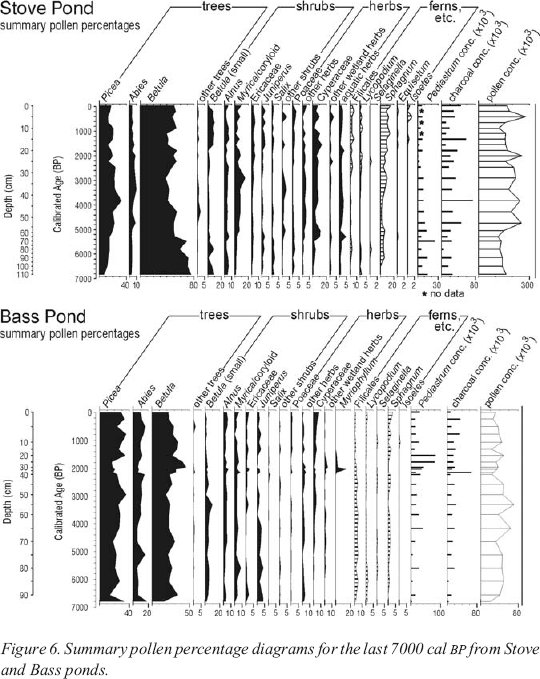
Pollen diagram: representation
17 Shortly after 7000 cal BP, when the record begins, more than 80% of the pollen accumulating in the sediment was Betula (birch), and birch pollen continues to dominate the pollen profile throughout. Birch is not the major tree in the forest; today it makes up only 7% of the merchantable forest of the Great Northern Peninsula (Macpherson 1995). Its strong over-representation is the result of high pollen productivity and wind dispersal, together with the presence of tree birches of less than merchantable size. The percentage of Picea (spruce) pollen (17%) is more closely related to the proportion of merchantable spruce trees in the vegetation (20%), while Abies (balsam fir) is greatly under-represented, less than 10% of the recent total pollen representing 65% of the merchantable forest. The insect-pollinated but widespread heaths (Ericaceae) are also under-represented.
Bog development
18 Based on the pollen assemblages, it appears that the composition of the forest at Stove Pond 7000 cal BP was not greatly different from the present, with somewhat less spruce and relatively more birch. At that time, however, the forest was probably more widespread than it is today, when it is confined to the well-drained slopes of the moraine east of the lake.
19 By 6200 cal BP there is evidence of the beginning of a major change in the landscape as a whole, marked by the increase of the spores of Sphagnum. These mosses grow on the floors of damp forests and the accumulation of their remains eventually prevents the regeneration of trees in the process of paludification (Bouchard et al. 1978; Davis 1984). Peat thicknesses may eventually reach several metres, placing the minerals in the underlying soil beyond the reach of plant roots. Those plants surviving on the bog surface are specialized, such as Sphagnum itself, or stunted.
20 Increasing peat cover on the mineral soil diminishes the input of mineral sediment to the lake, reducing the rate of sediment accumulation. In the case of Stove Pond this is apparent after 5000 cal BP, and explains the increasing pollen concentrations. In addition, the reduction of dissolved mineral input to the lake restricted the growth of aquatic organisms such as Pediastrum.
21 The onset of paludification varied in time across the island of Newfoundland, depending on the local moisture balance and topography. Davis (1984) found that bog inception was more frequent before 6800 cal BP and after 4500 cal BP; an interval of low Sphagnum values at Stove Pond coincides with the end of this period and appears to represent dryer and/or warmer conditions.
Development of vegetation and interpretation
22 Between 7000 cal BP and the present the pollen record indicates that there were changes in the composition of the forest. The proportion of spruce pollen increased after 7000 cal BP, reaching a peak at 3800 cal BP, coincident with the maximum charcoal concentration. Peak values for balsam fir pollen were registered from 4800 to 2000 cal BP, with a slight decrease around 4300 cal BP, coinciding with low values for Sphagnum spores. At that time values for the pollen of “other trees” (mainly Fraxinus (ash)) were at a maximum. It may be that ash grew closer to Stove Pond than it does today, when it grows no further north on the west coast of Newfoundland than Bonne Bay. Betula (birch) pollen percentages declined irregularly, registering relatively low values 3800-3000 cal BP, with a further interval of low values 1600-1100 cal BP.
23 All the shrubs and herbs represented in the pollen diagram can survive damp conditions, and some require them. Since the proportions of shrub and herb pollen and of fern spores fluctuate with that of Sphagnum, they appear to be related to the development of the bog. Of the herbs, Cyperaceae (sedges) would have grown first along streams, on “sedge meadows” (Bouchard et al. 1978) and at the edge of the lake; later these plants would have grown on the surface of the bog. The taxa grouped as “aquatic herbs” include Menyanthes (buckbean) and Utricularia (bladderwort), and require or can survive in shallow water. Poaceae (grasses) include species that can grow in damp situations and on bogs, as do the Rosaceae, the most frequent of the “other herbs.” Myrica gale (sweet gale) grows near water and on bogs; its representation decreases with increases in the pollen of, first, Ericaceae (heaths), followed by small (shrub) birch, Juniperus (common or creeping juniper) and finally Salix (willow), a sequence suggesting changing conditions on the bog surface. Dwarfed spruce and balsam fir trees may also have been present on the bog, or in patches of tuckamore, although much greater amounts of pollen would have been produced by the trees growing on the moraine to the east of the lake.
24 The spores of ferns (Filicales), clubmosses (Lycopodiaceae), spikemoss (Selaginella) and horsetails (Equisetum) were produced by plants growing in moist habitats in forest or tuckamore, or on wetland. Their record echoes that of Sphagnum; abundances of all are reduced in the interval 5000-3000 cal BP, when peak values for the pollen of spruce and more temperate trees are registered.
25 The greatest charcoal concentration is registered at 4000 cal BP. There is no pollen evidence to suggest that fire occurred in the catchment; however, fire occur-rence and frequency may be greater in intervals of higher growing season temperature. Therefore, the occurrence of fire is consistent with other evidence of greater summer warmth at this time. For instance, at Bass Pond, at the north coast of the Point Riche Peninsula, the chironomid-based temperature profile indicates maximum Holocene warmth around 5000 cal BP (Rosenberg et al. this volume); a lag in the response of terrestrial vegetation is to be expected.
BASS POND
26 Bass Pond is a small lake (~ 5 ha) lying at an elevation of 8 metres within 100 metres of the shore of Port au Choix Cove, on the north side of the Point Riche Peninsula (Figure 1). It was selected for its proximity to the Groswater Palaeoeskimo site of Phillip’s Garden East, which is located 500 metres to the west (Figure 2; Renouf 1993).
27 The lake’s 30-hectare catchment is underlain mainly by the dolomitic Port au Choix formation, the surface of which rises southward to the foot of a limestone cliff in the overlying Table Head formation (Bostock et al. 1983). The highest point is at Crow Head, about 45 metres above sea level (Figure 2). A small cave at the base of the cliff was found to contain human remains and artifacts of Groswater age (Brown 1988; Renouf 1999). Small streams drain the wooded slope below Crow Head, entering Bass Pond after passing through the bordering fen. A belt of scrub or heath lies between the lake and the low sea cliffs (Bouchard et al. 1978; Northlands Associates 1985).
28 The rather open woodland inland from the pond consists primarily of small balsam fir (Abies balsamea) with white spruce (Picea glauca); white birch (Betula papyrifera) and mountain ash or dogberry (Sorbus americana) are the only hard-wood trees. The large pollen of Abies is seldom carried far by the wind, and degradation of many balsam fir and spruce grains indicates that some pollen was carried to the lake by streams or slope-wash. Larch (Larix laricina) is present, but its frequently degraded pollen was not identified in the assemblage. Mosses form the ground cover, including Sphagnum on wet sites, and the herb layer includes ferns (Filicales) as well as a variety of flowering plants. Alder (Alnus) is not found in the present vegetation although Alnus pollen does occur in the record (Figure 6). The fen bordering the southern shore of the lake (Northland Associates 1985) supports a varied vegetation including sedges (Cyperaceae), heaths (Ericaceae), shrub birches, sweet gale (Myrica gale), dwarfed spruce, herbs and some Sphagnum.
29 The scrub between the lake and the cliff-top is dominated by heaths including Empetrum, together with dwarf birch, dwarfed balsam fir and black spruce (Picea mariana), juniper (Juniperus), sweet gale, willows (Salix) and sedges. Pollen from this area could have been carried to the pond by onshore winds.
30 A 2.1-metre sediment core was retrieved from a water depth of 0.94 centimetres from a central sediment-filled karst depression in July 1987. Except at the base, the sediment consisted of marl. A further core was retrieved in April 1993 to obtain material for radiocarbon dating (Table 1) and was matched with the first by pollen analysis. All of the radiocarbon dates, except for the basal sample, are on terrestrial macrofossils analyzed by atomic mass spectrometry (see Rosenberg et al. this volume for a discussion of radiocarbon dates from Bass Pond).
31 Macrofossils were most frequent at sediment depths of 22-29 centimetres and 40-60 centimetres, and consisted principally of twig fragments. Balsam fir was the most frequently identified species (P.J. Scott, pers. comm. 1996). Figure 6 is a summary pollen diagram from the upper part of the initial core. Sediment accumulated most rapidly between 2200 and 1000 cal BP (40-10 cm) and particularly between 2200 and 2000 cal BP (40-30 cm), accounting for the low pollen concentrations in this interval.
32 Pollen concentrations overall are much lower than at Stove Pond. The presence of Alnus pollen in proportions similar to those at Stove Pond, despite the current absence of alder from the catchment, indicates that a component of the pollen rain is wind-borne. To what extent, then, do the proportions of other wind-borne grains, especially Picea and Betula, represent the catchment’s vegetation? Perturbations in the pollen curves for Picea and Betula coincident with a sharp decline and subsequent increase in local Abies between 2200 and 1600 cal BP suggest that a significant proportion of all these grains is of local origin and that trends in these curves may match trends in the vegetation.
33 Before about 3000 cal BP the curves for the major taxa show few fluctuations, suggesting no marked changes in the vegetation of the catchment. At 3000 cal BP both Picea and Abies pollen increase at the expense of Betula, both “tree” (>20 μm) and “shrub” (<20 μm) pollen types. This suggests that a proportion of the grains classified as “tree” birch are in fact from shrubs. It follows that the conifer woodland in the Bass Pond catchment attained its greatest height and density at about 3000 cal BP.
34 Abies pollen maintained high values until 2200 cal BP, while Picea pollen declined in abundance. After 2200 cal BP a change is registered in many taxa, coincident with a peak of charcoal, and the perturbation continued until 1800 cal BP. Picea and Abies pollen declined rapidly, and then made a slow recovery. “Tree” birch pollen increased to a maximum and then declined. Pollen from most shrub and herb taxa show short-lived peaks in the 2200-2000 cal BP interval. However, Salix pollen decreased in abundance and Sphagnum spores were reduced to very low levels between 2200 and 1800 cal BP. Because these plants occur only close to the pond, there appears to have been disturbance of the vegetation at the pond shore, as well as more widely within the catchment.
35 Pollen from the aquatic herb Myriophyllum first appeared and peaked briefly at 2000 cal BP, and subsequently decreased in abundance and disappeared. This taxon is noted as occurring currently in some eutrophic (nutrient-rich) lakes within the region (Bouchard et al. 1978), and a rare species, Myriophyllum sibiricum, was recorded in Bass Pond in the early 1990s (M. Burzynski, pers. comm. 2004). The coincident increase in remains of the aquatic alga, Pediastrum (which continued until 1400 cal BP), also suggests increased nutrient availability. The fire indicated by the charcoal peak at 2200 cal BP may have released nutrients into the catchment soils and thence to the lake (Cwynar 1978; Burden et al. 1986), as would anthropogenic or other disturbances within the catchment.
36 The evidence for widespread vegetation change between about 2200 and 1800 cal BP, together with increased sediment deposition and fire, suggests considerable disturbance in the drainage basin. The lack of evidence for a correlative disturbance in Stove Pond points to a localized event on the Point Riche Peninsula. Even after 1500 cal BP, when pollen spectra largely resume their pre-disturbance values, fluctuations in many taxa suggest further but less marked disturbance. Low abundances of balsam fir and juniper pollen and of Sphagnum spores, together with increased abundance of (shrub?) birch pollen at 1100 cal BP, indicate one disturbance episode. At 500 cal BP low spruce pollen abundance is accompanied by increases in pollen from balsam fir, birch and other trees, a decline in juniper and very low values for Lycopodium and Sphagnum. Because balsam fir, dwarfed spruce, juniper, clubmoss, and Sphagnum together represent the vegetation surrounding the lake and the coastal heath, these disturbances are interpreted as being confined largely to the lake shore and coastal cliff top. The apparently low rate of accumulation of the top 10 centimetres of sediment precludes finer temporal resolution of this part of the record.
HUMAN IMPACT ON THE ENVIRONMENT
37 The clearest evidence for disturbance in the Bass Pond drainage basin appears at 2200 cal BP and is sustained for at least 400 years until 1800 cal BP. This coincides with the end of the Groswater Palaeoeskimo occupation of the Phillip’s Garden headland (2950-1820 cal BP) and the beginning of the Dorset occupation of Phillip’s Garden (1990-1180 cal BP). Keeping in mind that radiocarbon dating is imprecise and that temporal coincidence does not necessarily mean causality, we nevertheless suggest three possible connections between the Bass Pond disturbance and human activities in its immediate vicinity. First, we suggest that the late Groswater occupation may have caused the 2200 BP disturbance, primarily through fire, but also through tree cutting and wood-working. Second, we suggest that people modified the pond-edge vegetation by trampling during the Groswater and Dorset occupations, spanning 2200 and 1800 cal BP. Third, we suggest that increased nutrient input in the pond from 2000 cal BP onward might have been the result of Dorset sealskin processing.
38 Dealing with each in turn, it is possible that the Bass Pond disturbance was triggered by a forest fire at 2200 cal BP, as indicated by the dramatic peak in charcoal at this time. This could have led to associated changes in vegetation, including the marked decline in Picea and Abies and a corresponding increase in shrubs and herbs. This would have been a local rather than a regional forest fire, since there is no record of it in Stove Pond. Although the fire could have been natural, the modern-day occurrence of fire on these northern coastal barrens is extremely rare (Meades and Moores 1989), and the charcoal peak occurred during a period of particular cold, indicated by summer water temperatures in Bass Pond (Rosenberg et al. this volume). An alternative possibility is that this forest fire was started by the Groswater people who occupied the area. This likely would have been accidental, because seal hunters would derive no particular benefit from the fire, unlike terrestrial-based hunter-gatherers elsewhere who were known to set fires to attract herbivores to the new post-fire vegetation growth (Cummins 2000; Williams 2002). Either the hearths or the heated rocks, the main source of heat for the Groswater people, could have started a local fire.
39 Wood-cutting or wood-working may be indicated by the four axes from Phillip’s Garden East and one small gouge that was found at Phillip’s Garden West. Wood-working might have contributed to the decrease in Picea and Abies. This activity might have produced the distinct band of twig fragments, mostly balsam fir, at the 22-29-centimetre level that dates to approximately 2400-2000 cal BP.
40 Lakeside disturbance is also reflected in a marked reduction in the occurrence of Salix and Sphagnum between 2200 and 1800 cal BP. Although the early period of this reduction might be linked to the fire, its persistence over the longer term suggests other contributing factors. One possibility is trampling first by Groswater and later by Dorset people, as they engaged in pond-side activities. Sphagnum, which is particularly vulnerable to trampling, does not return to pre-disturbance levels until 1400 cal BP.
41 There are elevated nutrient levels in Bass Pond at 2200-1800 cal BP, indicated by increased concentrations of Myriophyllum and Pediastrum, the former peaking at 2000 cal BP. There is also a sharp increase in sedimentation at 2200-1100 cal BP. Initially this might have been a response to soil erosion in the drainage basin following the fire disturbance at 2200 cal BP; however, the sustained nature of the disturbance suggests a more complex situation. Although shrub and herb taxa returned to normal levels by 2000 cal BP, tree taxa do not show substantial recovery until 1500 cal BP, Pediastrum reached a maximum concentration at 1900 cal BP and remained high until about 1400 cal BP when pre-disturbance levels were abruptly re-established. Sedimentation rates remained high until 1100 cal BP. This continued disturbance, particularly the late peak and persistence of high Pediastrum concentrations and the increased sedimentation rate, may be related to activities of Dorset people in the area following their occupation of Phillip’s Garden at 1900 cal BP.
42 In addition, there is an intriguing temporal correspondence between high salinity in Bass Pond and the beginning of Dorset occupation. Fossil chironomid assemblages in the Bass Pond sediments (Rosenberg et al. this volume) indicate that at 2000 cal BP there was a dramatic rise in salinity to 1.9 grams/litre. This salinity peak persisted until 1600 cal BP and freshwater conditions returned to Bass Pond about 1100 cal BP, which corresponds to the abandonment of Phillip’s Garden by the Dorset. Rosenberg et al. (this volume) speculate that a marine incursion, or the reworking of marine sediments in the drainage basin may have been responsible for this salinity peak. Alternatively, an activity that might be associated with the elevated nutrient status and salinity of Bass Pond and the increased rate of sedimentation, is sealskin processing, in particular for the production of leather. We have already suggested the possibility of sealskin processing for the Groswater occupation. However, given the increased size of the Phillip’s Garden site, and evidence for more intensity of site use with an almost exclusive focus on harp seal, sealskin processing might have been at a larger scale and have left its environmental mark.
43 Throughout the Arctic, sealskin is prized for its water-proof properties and is an important raw material for tent covers, parkas and footwear (Balikci 1970). Abundant faunal material from Phillip’s Garden demonstrates that the prime economic purpose of this large and intensively occupied site was the harp seal hunt. It is highly likely that in addition to hunting seal for meat and fat, the Phillip’s Garden Dorset processed the hides for garments and boots. Based on the ethnographic descriptions of Inuit sealskin processing (Oakes and Riewe 1996) and sealskin processing on the Great Northern Peninsula (Multimedia Creations 2002), several sealskin processing tools can be identified, including bone awls, tabular slate scrapers and chert scapers. Bone awls would have been used to punch holes in the skins for lashing them to a frame. Tabular scrapers were specialized for removing the fat from the hide (Gracie 2004) (Figure 7), and were the Dorset version of the ulu. The multi-purpose chert end-scraper could also have been used to scrape the fat from the skin, although likely after the initial and primary fat removal using the larger slate scraper.
Figure 7. An example of a tabular slate tool from Phillip’s Garden, interpreted as sealskin scraping tools. The dime for scale is 1.7 cm in diameter.

Display large image of Figure 7
44 A recent detailed description of sealskin curing on the northern part, or Strait coast, of the Great Northern Peninsula, provides contextual background for understanding the archaeological record. In this area, sealskin curing has a long tradition thought to be borrowed from the Labrador Inuit (Firestone 1992, 1994). As described by Bock (1991) and shown in detail in Multimedia Creations Inc. (2002), after the seal harvest, pelts were separated from the seal carcass and rolled in salt to preserve them until early summer. At that time as much as possible of the fat from the inside of the pelt was removed using a curved knife, and the pelt was stretched out on a wooden frame and placed in the sun to dry. Next was a dry scrape with a metal scraper to remove still more fat; this tool was set in a handle so that the working edge was at right angles to the hide. This tool is larger than, but similar in working edge to the Dorset chert scraper. Of particular interest to the discussion of the rise in salinity in Bass Pond, the framed pelt was then placed in a freshwater pond (Figure 8) and bacterial agents effectively “de-haired” the pelt, a natural process which usually took three weeks. The last stage involved bark-tanning the pelt for five to seven days in a salt-water solution of rinds (bark) from spruce, fir and birch trees, before the skins were removed to dry.
Figure 8. Photograph of a sealskin (110 by 90 cm) submerged in a shallow freshwater pond to be de-haired by natural bacterial agents; near Green Island Cove, on the northwestern coast of the Great Northern Peninsula.

Display large image of Figure 8
45 While the Dorset would not have had access to salt to preserve the skins, the salt content on the skin from its original seawater environment could be enough to affect the salinity of Bass Pond, especially if a large number of sealskins were processed at the one time. Or, more likely, if they tanned the skins at the location of processing, Bass Pond, when finished they likely would have tipped the salt-water and rind solution into the pond. Meanwhile, the decay that accompanied the removal of hair would have increased lake nutrient status, perhaps leading to an increase in amounts of the aquatic herb Myriophyllum and the aquatic alga Pediastrum.
46 On this basis we propose that pelt curing activities in and near Bass Pond may have led to eutrophication of the lake water between 1900 and 1400 cal BP. Similar conclusions about human disturbance in pond sediments were reached by Douglas et al. (2004) who documented elevated nutrient concentrations in a pond beside a Thule Inuit whale butchering site (AD 1600-1200) on Somerset Island.
CONCLUSIONS
47 Macro- and microfossil data from Bass Pond, Port au Choix, demonstrate that at 2200-1800 cal BP there were marked changes in the local environment, in particular an abrupt decrease in the local spruce and fir forest, likely due to an intense and rare forest fire; a persistent disturbance of pond-side vegetation; and salinization and eutrophication of lake waters. We show that these changes were coincident with adjacent Groswater and Dorset Palaeoeskimo occupations, and argue that their activities, such as tree cutting, wood burning and working, pond-side trampling, and seal skin processing, were related to these disturbances. Further lake sediment analysis, including diatoms and sediment geochemistry, is planned to refine the nature and character of the disturbance events.
Acknowledgements
Multi-year funding has been provided by the Natural Sciences and Engineering Research Council of Canada, the Social Sciences and Humanities Research Council of Canada, Parks Canada, and Memorial University of Newfoundland. Dr. Peter Scott, Biology, Memorial University, identified macrofossils. Diagrams were drafted by David Mercer, Memorial University of Newfoundland Cartography Laboratory. Comments by two reviewers are gratefully acknowledged.References
Aronsson, K.-Å. (1994), “Pollen evidence of Saami settlement and reindeer herding in the boreal forest of northernmost Sweden — an example of modern pollen rain studies as an aid in the interpretation of marginal human interference from fossil pollen data.” Review of Palaeobotany and Palynology 82: 37-45.
Balikci, A. (1970), The Netsilik Eskimo. New York: Natural History Press.
Behre, K.-E., ed. (1986), Anthropogenic indicators in pollen diagrams. Rotterdam: Balkema, 232 p.
Bock, A. (1991), Out of Necessity: the Story of Sealskin Boots in the Strait of Belle Isle. Great Northern Peninsula Craft Producers. Shoal Cove, NL.
Bostock, H.H., L.M. Cumming, H.H. Williams, and W.R. Smyth (1983), Geology of the Strait of Belle Isle Area, Northwestern Insular Newfoundland, Southern Labrador, and adjacent Quebec. Geological Survey of Canada, Memoir 400, 145 p. and maps.
Bouchard, A., S. Hay, and E. Rouleau (1978), “The vascular flora of St. Barbe South District, Newfoundland: an interpretation based on biophysiographic areas.” Rhodora 80: 228-308.
Brown, S.C. (1988), Archaeological Investigations at Crow Head Cave and the Gargamelle Rock Shelter in the Port au Choix National Historic Park, Newfoundland. Ms. on file, Parks Canada, Atlantic Region, Halifax.
Burden, E.T., J.H. McAndrews, and G. Norris (1986), “Palynology of Indian and European forest clearance and farming in lake sediment cores from Awenda Provincial Park, Ontario.” Canadian Journal of Earth Sciences 23: 43-54.
Cummins, G. (2000), “Fire! Accidental or Strategic Use of Fire in the Early Mesolithic of the Eastern Vale of Pickering.” In Mesolithic Lifeways: Current Research from Britain and Ireland, edited by R. Young, pp. 74-84.
Leicester Archaeological Monographs,7. School of Archaeological Studies, Leicester.
Cwynar, L.C. (1978), “Recent history of fire and vegetation from laminated sediment of Greenleaf Lake, Algonquin Park, Ontario.” Canadian Journal of Botany 56: 10-21.
Davis, A.M. (1984), “Ombrotrophic peatlands in Newfoundland, Canada: their origins, development and trans-Atlantic affinities.” Chemical Geology 44: 287-309.
Douglas, M.S.V., J.P. Smol, J.M. Savelle, and J.M. Blais (2004), “Prehistoric Inuit Whalers Affected Arctic Freshwater Ecosystems.” PNAS 101: 1613-1617.
Erwin, J.C. (1995), “An Intra-site Analysis of Phillip’s Garden: A Middle Dorset Palaeo-Eskimo Site at Port au Choix, Newfoundland.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
Firestone, M. (1992), “Inuit Derived Culture Traits in Northern Newfoundland.” Arctic Anthropology 29 (1): 112-128.
Firestone, M. (1994), “Skin Boots, Coach Boxes and the Ski-doo: The Renaissance of Traditional Items in Northern Newfoundland.” Material Culture 26 (2): 15-27.
Gracie, N. (2004), “Function and Manufacture of Tabular Ground Slate Tools from Phillip’s Garden, Newfoundland.” Hons. thesis, Department of Anthropology, Memorial University of Newfoundland.
Grant, D.R. (1994), Quaternary geology of Port Saunders map area, Newfoundland. Geological Survey of Canada, Paper 91-20, 59 p.
Harp, E. (1976), “Dorset Settlement Patterns in Newfoundland and Southeastern Hudson Bay.” In Eastern Arctic Prehistory: Palaeoeskimo Problems, edited by M.M. Maxwell, pp. 119-138. Washington: Memoir for the Society for American Archaeology.
Hicks, S. (1993), “Pollen evidence of localized impact on the vegetation of northernmost Finland by hunter-gatherers.” Veget. Hist. Archaeobot 2: 137-144.
Hodgetts, L.M. (this volume), “Using Bone Measurements to Determine the Season of HarpSeal Hunting at Phillip’s Garden.”
Hodgetts, L.M., M.A.P. Renouf, M.S. Murray, L. Howse, and D. Balkwill (2003), “Changing Diet Breadth at the Dorset Palaeoeskimo Site of Phillip’s Garden, Newfoundland.” Arctic Anthropology 40 (1): 106-120.
Kennett, B.L. (1990), “Phillip’s Garden East: A New Perspective on Groswater Palaeoeskimos.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
LeBlanc, S. (1996), “A Place with a View: Groswater Subsistence-Settlement Patterns in the Gulf of St. Lawrence.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
Macpherson, J.B. (1995), “A 6ka BP reconstruction for the island of Newfoundland from a synthesis of Holocene lake-sediment pollen records.” Géographie physique et Quaternaire 49 (1): 163-182.
Macpherson, J.B. (1997), “Stop 20: Port au Choix Palaeoenvironments.” In Great Northern Peninsula and Southern Labrador Field Trip Guide by T. Bell, D.G.L. Liverman, S. Evans, and M. Batterson, prepared for the 1997 meeting of the Canadian Association of Geographers, Department of Geography, Memorial University of Newfoundland, 92 p.
Meades, W.J. and L. Moores (1989), Forest site classification manual: a field guide to the Damman forest types of Newfoundland. Ministry of Supply and Services Canada, and Newfoundland Department of Forestry and Agriculture.
Multimedia Creations Inc. (2002), Sealskin Craft Production and Bark-Tanning. CD produced in partnership with the Great Northern Peninsula Craft Producers. Shoal Cove, Newfoundland.
Murray, M.S. (1992), “Beyond the Laundry List: The Analysis of Faunal Remains from a Dorset Dwelling at Phillip’s Garden (EeBi 1), Port au Choix, Newfoundland.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
Northland Associates Ltd., St. John’s (1985), Natural Resources Inventory, Port au Choix National Historic Park. Parks Canada Contract No. 84-HSN-021.
Oakes, J. and R. Riewe (1996), Our Boots: an Inuit Women’s Art, New York: Thames andHudson.
Rankin, C. and Y. Labreche (1991), “Traditional Ulus and their Prehistoric Counterparts in the Central and Eastern Canadian Arctic.” Etudes/Inuit/Studies 15 (1): 105-130.
Renouf, M.A.P. (1993), “Palaeoeskimo seal hunters at Port au Choix, northwestern Newfoundland.” Newfoundland Studies 9 (2): 185-212.
Renouf, M.A.P. (1994), “Two Transitional Sites at Port au Choix, Northwestern Newfoundland.” In Threads of Arctic Prehistory: Papers in Honour of William E. Taylor, Jr., edited by D. Morrison and J.-L. Pilon, pp. 165-196. Archaeological Survey of Canada, Mercury Series 149. Hull, Quebec: Canadian Museum of Civilization.
Renouf, M.A.P. (1999), Ancient Cultures, Bountiful Seas: the Story of Port au Choix. St. John’s: Historic Sites Association of Newfoundland and Labrador.
Renouf, M.A.P. (2002), Archaeology at Port au Choix: 1990-1992 Excavations. OccasionalPapers in Northeastern Archaeology, 12. St. John’s: Copetown Press.
Renouf, M.A.P. (2004), Re-investigating a Middle Phase Dorset Dwelling at Phillip’s Garden, Port au Choix, Newfoundland. Proceedings of the SILA/NABO conference, edited by B. Gronnow. Copenhagen: The Danish National Museum (accepted for publication).
Rosenberg, S.M., I.R. Walker, and J.B. Macpherson (this volume), “Environmental Changes at Port au Choix as Reconstructed from Fossil Midges.”
Ryan, A.G. (1978), Native Trees and Shrubs of Newfoundland and Labrador. Parks Division, Department of Tourism, Government of Newfoundland and Labrador, 120 p.
Sergeant, D. (1991), Harp Seals, Man and Ice. Canadian Special Publications of Fisheries and Aquatic Sciences 114. Ottawa: Department of Fisheries and Oceans.
Stuiver, M. and P.J. Reimer (1993), “Extended 14C Database and Revised CALIB Radiocarbon Calibration Program.” Radiocarbon 35: 215-230.
Wells, P.J. (this volume), “Animal Exploitation and Season of Occupation at the GroswaterPalaeoeskimo Site of Phillip’s Garden West.”
Wells, P.J. (2002), “An Analysis of Faunal Remains from Two Groswater Palaeoeskimo Sites at Port au Choix, Northwestern Newfoundland: Phillip’s Garden West (EeBi-11) and Phillip’s Garden East (EeBi-1).” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
Williams, G.W. (2002), “Aboriginal Use of Fire: Are there any “natural” plant communities?” In Wilderness and Political Ecology; Aboriginal Influences and the Original State of Nature, edited by Charles E. Kay and Randy T. Simmons, pp. 179-211. Salt Lake City: The University of Utah Press.
Notes