Using Bone Measurements to Determine the Season of Harp Seal Hunting at the Dorset Palaeoeskimo Site of Phillip’s Garden
Lisa M. HodgettsDepartment of Anthropology at the University of Western Ontario
INTRODUCTION
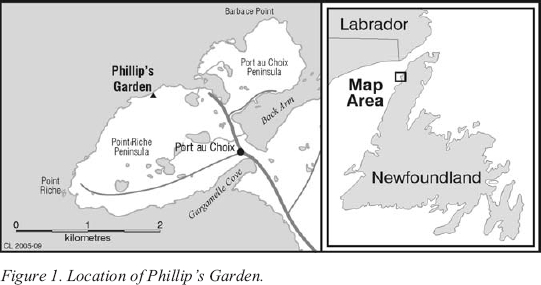
1 THE DORSET PALAEOESKIMO were arctic-adapted hunter-gatherers who occupied the Canadian arctic, Greenland, the Labrador coast, and Newfoundland between about 2500 and 800 radiocarbon years ago. While there are several known Dorset sites in the Port au Choix region, the most extensively excavated is Phillip’s Garden, located on the outer coast of the Point Riche Peninsula. It is the largest known Dorset site in Newfoundland, with the depressions from over 60 semi-subterranean dwellings visible on the ground surface (Figure 1). The site was occupied from approximately 1990 to 1180 cal BP.1 Archaeological excavations at the site have revealed large waste deposits, or middens, that contain vast quantities of animal bone discarded by its Dorset occupants during food preparation and consumption (Harp 1976; Renouf 1986a, 1987, 1991, 1993; Hodgetts 2002). Harp seal remains dominate these collections of bone (cf. Harp 1976; Renouf and Murray 1999; Renouf 2000, 2002), indicating that activities at the site focused intensively on harp seal hunting.
2 Today, harp seals ( Phoca groenlandica) migrate through the Strait of Belle Isle twice each year on their journey from their summer feeding grounds in arctic waters to their breeding grounds in the Gulf of St. Lawrence. Harp seals feed at the ice edge but require stable pack ice on which to give birth to their young (Sergeant 1991). Their annual movements have been attributed to seasonal variations in ice conditions and the availability and level of competition for favoured prey species such as capelin (Sergeant 1991).
Figure 1. Location of Phillip’s Garden.
Display large image of Figure 1
3 As mobile hunter-gatherers, the Dorset moved around the landscape in order to exploit seasonally available resources. There are few faunal assemblages from Dorset sites in Newfoundland, and as a result there is limited direct evidence of Dorset diet in the region. Archaeologists have used information about site location to create a tentative picture of Dorset subsistence and settlement patterns in Newfoundland. Schwarz (1994) provides the most recent synthesis. He proposes that the Dorset spent the spring at the outer coast hunting harp seals, and summered at inner coastal locations where they exploited a range of birds, seals, land mammals, and inshore fish. Few Dorset sites are known in the interior, but they may have moved inland in the autumn to hunt caribou. Their winter activities remain elusive. Schwarz (1994: 65) suggests that they may have returned to the outer coast in December, to hunt harp seals during their southward migration. He also raises the possibility (Schwarz 1994: 66) of an extended winter harp seal hunt on the northeast coast of Newfoundland based on nineteenth-century accounts of successful winter harp seal hunting in this area under certain wind conditions. Schwarz maintains that the Dorset adaptation in Newfoundland was heavily reliant on harp seal, with potentially the entire winter and spring devoted to harp seal hunting at the outer coast. This model, however, is something of an oversimplification. Like any human group, the Dorset in Newfoundland were neither static nor entirely homogeneous. Their settlement and subsistence practices no doubt varied over time, both from year to year during the life of any particular individual, and on a broad scale from century to century (cf. Hodgetts et al. 2003). Regional variations have also been demonstrated in Newfoundland Dorset settlement patterns (Robbins 1985) and stone tool styles (LeBlanc 1999). There were doubtless similar regional variations in subsistence strategies, given that the availability of most prey species varies across the island.
4 Phillip’s Garden represents one element of a larger Dorset settlement and subsistence system. Despite extensive archaeological work on the site, the precise nature of its role within this system is only partially understood. The site was obviously used for hunting seals, and its large size suggests that it may have been home to seasonal aggregations of the Dorset population in the region (Renouf 2000). The season or seasons of occupation at Phillip’s Garden remain the subject of speculation. Spring hunting has been demonstrated at the site based on the presence of newborn seal bones (Renouf and Murray 1999). However, the migratory movements of harp seals in the area suggest the possibility of a second hunt in early winter (December-January). Until recently, this early winter hunt could not be identified based on the archaeological evidence. It can now be conclusively demonstrated through a comparison of measurements taken on seal bones from the site, with those taken on bones of modern harp seals with a known season of death.
MODERN HARP SEAL MIGRATION PATTERNS
5 The northwest Atlantic population of harp seals breeds in the waters off Newfoundland and Labrador (Figure 2). The following description of their migration patterns is taken from Sergeant (1991: 33-56) and LeBlanc (1996: 24-28; 2000). Early each fall, the seals leave their summer feeding grounds among the Arctic islands and move southward along the Labrador coast. They reach the Strait of Belle Isle around mid-December. Here they divide into two main breeding populations, the Front herd and the Gulf herd. The Front herd remains off the coast of Labrador and northeastern Newfoundland while the Gulf herd passes through the Strait of Belle Isle and continues southward to the Gulf of St. Lawrence. A smaller, poorly understood breeding population known as the Mecatina patch also forms on occasion off the Quebec lower north shore.
6 As the Gulf herd is moving south towards its breeding grounds, individuals tend to remain within a few kilometres of the Labrador/Quebec shore (LePage 1989), though in some years they are also found close to Newfoundland (Renouf 1986b). The Gulf herd gathers on the ice in late February and March to whelp and mate. Whelping groups regularly form near the Magdalen Islands, where the thickest ice is found (Sergeant 1991: 37-40). The seals then disperse briefly to feed. They congregate on the ice again in April to moult, a process that lasts approximately four weeks. In May they begin their migration to the north. Some individuals travel east initially, moving north along the east coast of Newfoundland, others take a more direct route up the west coast of Newfoundland (Figure 2).
7 Harp seal whelping and moulting grounds are generally located far out at sea on constantly shifting pack ice. This poses no obstacle to modern hunters equipped with large boats. However, it would have been difficult for the Dorset, hunting on foot or in small kayak-type boats, to reach the seals at this time. Instead, they probably hunted harp seals when the animals could be found closer to shore during their migrations (LeBlanc 1996: 34-38; 2000). Harp seal migration routes appear to be very stable, despite long-term climatic variation,2 and are unlikely to have undergone significant change over the past 2000 years. The Dorset at Phillip’s Garden were therefore presented with two options for hunting harp seal: an early winter hunt during the southward migration and a spring hunt during the northward migration. Outside their breeding season, harp seals live at the ice edge. The precise timing of harp seal migrations and the movements of individual harp seals are therefore driven largely by seasonal shifts in the location of sea ice (Sergeant 1991). Changing climatic conditions over the course of the Dorset occupation at Phillip’s Garden would have affected the distribution and extent of sea ice in the Gulf of St. Lawrence, causing variation in the availability of harp seals at the site, and in the exact timing and length of the hunting seasons.
Figure 2. Migration routes and breeding areas of the northwest Atlantic harp seal population.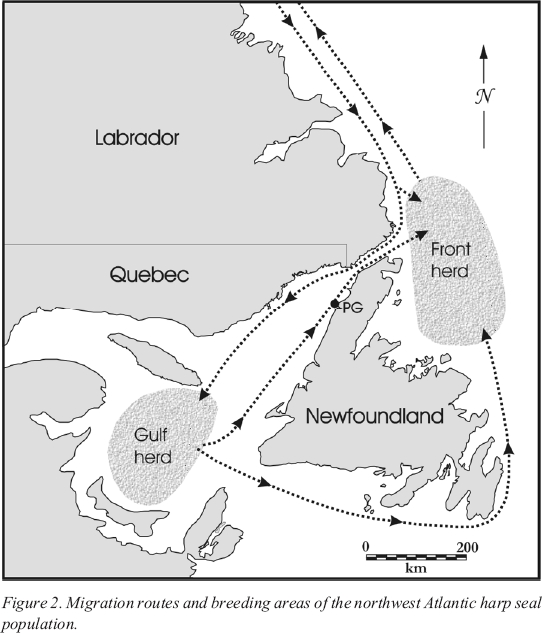
SEASONAL INDICATORS AT PHILLIP’S GARDEN
8 A precise seasonal determination for the harp seal hunt at Port au Choix could contribute to our understanding of the use of the site, and its role in Dorset settlement patterns on the west coast of Newfoundland. Spring harp seal hunting is indicated at Phillip’s Garden by the presence of a small number of newborn bones among the seal remains. Newborn bones are easy to identify based on their small size and spongy surface texture.
9 The early winter hunt is more difficult to establish. The bones of three-month-old juvenile harp seals (killed during the spring migration) and ten-month-old juveniles (killed during the early winter migration) cannot be separated on the basis of texture and shape alone. The early winter migration therefore cannot be conclusively demonstrated on the basis of a visual inspection of the seal remains. Several migratory bird species that winter in Newfoundland have been identified in the faunal assemblages from Phillip’s Garden. These include king eider, white-winged scoter, and dovekie. However, the presence of these winter migrants does not provide conclusive evidence of winter activity at the site, because many of these species are still found in the area in the spring (Table 1). Moreover, each of these species is found in very small numbers. In many cases they are represented by only single fragments from a limited number of features. It would therefore be unwise to use them as a basis for seasonal interpretation.
10 Seal teeth can act as an excellent indicator of season of death, since rates of tooth growth are not consistent throughout the year (cf. Bowen et al. 1983; Weber et al. 1993). In harp seals, growth is rapid during summer and early fall when the animals feed heavily. It slows in winter when food is less readily available, and in spring when the animals fast during their moulting period. This produces annual bands of tooth dentine, much like tree rings, which are visible in cross-section. Archaeological tooth sections can therefore be used to estimate season of death by comparing them to modern teeth from animals with known season of death. Unfortunately, attempts to use harp seal tooth sections from Phillip’s Garden to determine season of death have been unsuccessful (Hiseler 1997). Despite the excellent bone preservation exhibited at the site, the majority of teeth are in poor condition. The most recent growth layer, where seasonal information is read, was poorly defined in most of the sections. Of 58 canines that were sectioned, only four could be attributed to a particular season of death, all of them between March and June (Hiseler 1997: 114-128).
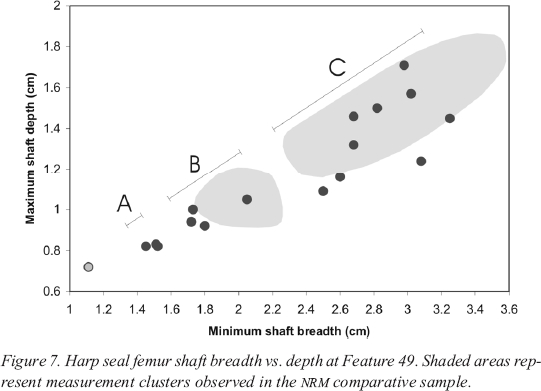
Table 1. Winter migrants present at Philip's Garden (sesonal information from Bull and Farrand 1994)

A MEASUREMENT TECHNIQUE FOR EXAMINING SEASON OF DEATH IN HARP SEALS
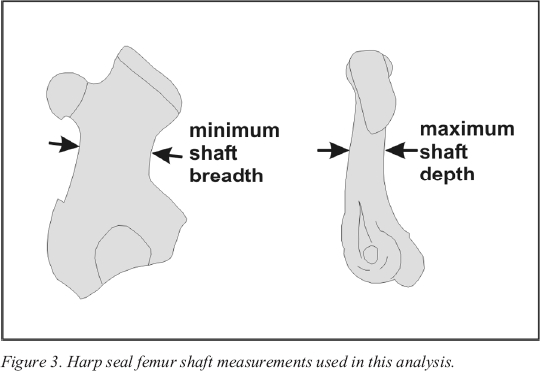
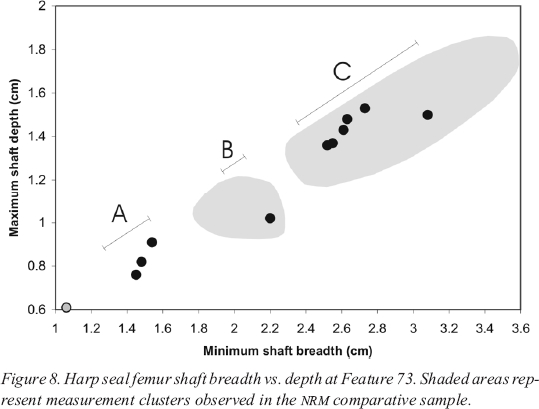
11 Limb bone measurements have been used effectively at several Stone Age sites in Scandinavia to determine the precise season of death of archaeological harp seal populations (Hodgetts 1999, 2001; Storå 1994, 2002). The technique relies on the fact that harp seals do not display marked sexual dimorphism; in younger individuals, males and females are roughly the same size at all stages of development, and adult males are only slightly larger on average than adult females (Maxwell 1967). Since harp seal pups are born within a short period each year, there are distinct age cohorts in the living population at any given time. These cohorts are reflected in the distribution of limb bone measurements from a seasonally hunted harp seal population. Storå (1994) has shown that a comparison of minimum femur shaft breadth and maximum femur shaft depth (Figure 3) is a particularly effective seasonal indicator.
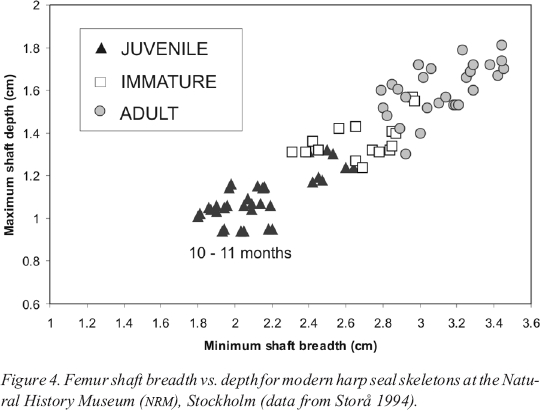
12 Figure 4 shows femur shaft breadth vs. depth for a modern collection of harp seals, housed at the Natural History Museum ( Naturhistoriska Riksmuseet — NRM) in Stockholm. These individuals were collected in January and February of 1987 in Tromsø, Norway. They belong to the White Sea population, where births peak in early March (Maxwell 1967). The measurement cluster in the lower left of the graph represents both male and female juveniles born the previous spring, making them ten to eleven months old when they were killed.3 To the right of these, and separated by a gap in the measurements, are the immatures. The youngest of these would have been approximately 22 months old. The eight anomalous “juveniles” that fall within the immature measurements are actually immatures, which were aged incorrectly when they were originally collected (Jan Storå pers. comm.). The immature femora grade in size into those of the adults. At Phillip’s Garden a spring hunt would therefore appear as two distinct clusters of measurements: two- to three-month-old juveniles on one hand, immatures and adults on the other. The addition of the December-January hunt would add a third central cluster of approximately ten-month-old juveniles.
Figure 3. Harp seal femur shaft measurements used in this analysis.
Display large image of Figure 3
Display large image of Figure 4
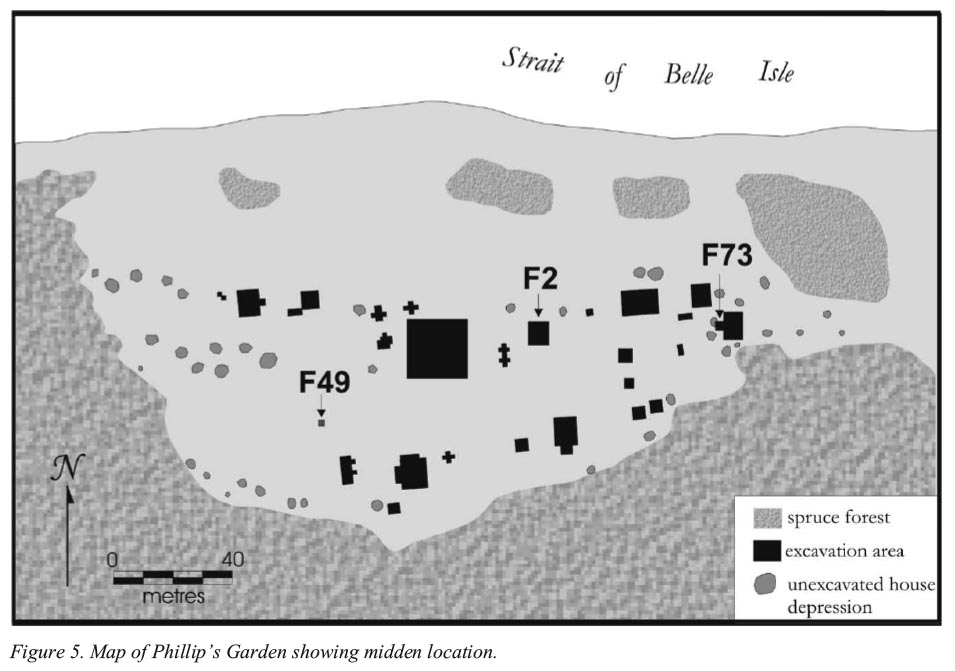
13 Faunal samples from three different middens, representing different stages in the occupation of the site, have been examined (Figure 5). The first of these, Feature 2, appears to have had at least two phases of use. An early phase is indicated by two dates, 1990-1870 cal BP (Beta 23977) and 1999-1710 cal BP (Beta 23978). A later phase of occupation is indicated by dates of 1530-1390 cal BP (Beta 15381) and 1520-1340 cal BP (Beta 19084). A second midden, Feature 49, produced a radiocarbon date of 1710-1920 cal BP (Beta 42967), which places it very early in the sequence of dates from the site. Finally, midden Feature 73 is one of the latest features on the site, with dates of 1400-1330 cal BP (Beta 160976), 1370-1180 cal BP (Beta 66436) and 1180-1350 cal BP (Beta 160977). The original radiocarbon dates are from Renouf (2002) and Hodgetts (2002).
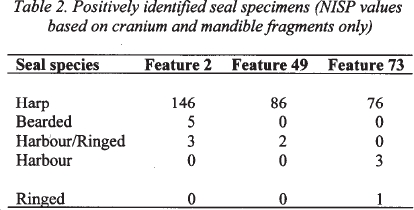
14 While the bones of the seal skull, and to a lesser degree the humerus, can be easily identified to species, other elements of the skeleton display a large degree of variation within each species, making them more difficult to identify. In addition to the harp seal, five other seal species are found in the Gulf of St. Lawrence: bearded seal ( Erignathus barbatus), hooded seal ( Cystophora cristata), grey seal ( Halichoerus grypus), harbour seal ( Phoca vitulina), and ringed seal ( Phoca hispida). Harp seal femurs can be readily distinguished from those of bearded seal, grey seal and ringed seal based on their shape (Hodgetts 1999, Appendix B). There is, however, some overlap in the range of femur size and shape observed in harp seals and that of harbour seals and hooded seals (particularly females). However, the dominance of harp seal crania and mandible fragments over those of all other seal species in the middens being examined (Table 2) means that “questionable” femurs could be attributed to harp seal with a high level of confidence.
15 Figure 6 illustrates the distribution of measurements taken on harp seal femora from midden Feature 2. The grey areas indicate the location of the two measurement clusters seen in the modern comparative sample from the Natural History Museum (NRM) in Stockholm. Many of the data points correspond closely with the two groups of measurements observed in the NRM comparative sample. The Group C measurements in the upper right of the graph overlap with, or lie just below (i.e., they display slightly smaller maximum shaft depths) the immature and adult measurements from the NRM. These can be safely interpreted as immature and adult individuals within the archaeological population. It is interesting to note that there is greater size variation within the archaeological material than the modern population. Unlike the reference sample, which was collected over a period of two months, the archaeological sample represents a composite of many years, or even decades, of hunting activity. The Feature 2 sample may therefore illustrate more of the diversity within the living population it represents than the NRM sample does. Alternatively, the Newfoundland harp seal population may simply exhibit a greater size range than the White Sea population.
16 The Group B measurements from Feature 2 correspond with the juvenile distribution from the NRM. Three specimens fall within the size range of the NRM juveniles and two lie just to the left. All five probably represent older juveniles killed during their southward migration in early winter. Today juveniles are approximately ten months old when they pass through the Strait of Belle Isle on their return to the Gulf of St. Lawrence. The juveniles from the NRM sample are slightly older on average, which may explain why two of the Feature 2 measurements in Group B fall slightly below the NRM distribution.
Figure 5. Map of Philip's Garden showing midden location.
Display large image of Figure 5
Table 2. Positively identified seal specimens (NISP values based on cranium and mandible fragments only)
Display large image of Table 2

Display large image of Figure 6
17 Group A, consisting of eight femora, is separated from the early winter juveniles in Group B by a marked break in the measurement distribution. The Group A specimens undoubtedly represent younger juveniles. Their small size suggests that they were killed during their spring migration to the north. Today, juveniles are approximately two to three months old at this time.
18 The measurement data from Feature 49 (Figure 7) look very similar to those from Feature 2 (Figure 6). The three groupings of measurements observed at Feature 2 are also found at Feature 49. Again, the measurements in Group C represent adults and immatures. They correspond nicely with the NRM reference collection measurements, (again indicated by shaded grey areas) for individuals over 22 months of age. As at Feature 2, there are two groupings of juvenile measurements. The larger juveniles of Group B are of similar size to the ten- to eleven-month old juveniles from the NRM sample. Early winter hunting at Feature 49 is represented by a single specimen that overlaps in size with the NRM juveniles, and three additional specimens that fall slightly below the NRM distribution. The data points in Group A indicate the presence of younger juveniles. As at Feature 2, there is a marked size difference between these smaller juveniles (Group A) and the larger winter killed ones (Group B). Here, too, the smaller juveniles indicate a spring hunt.
Figure 7. Harp seal femur shaft breadth vs. depth at Feature 49. Shaded areas represent measurement clusters observed in the NRM comparative sample.
Display large image of Figure 7
Display large image of Figure 8
19 The single grey data point in the lower left-hand corner of this graph represents a bone that was identified as foetal or neonatal based on its spongy surface texture. Foetal bone could indicate either a pregnant female killed anytime between the early winter southward migration and the spring whelping season, or a newborn pup killed in the spring. In the absence of a reference collection of foetal and neonatal bone from individuals at known stages of development, this class of bone will be excluded from the seasonal analysis.
20 Like the other two middens, Feature 73 contains three distinct clusters of femur measurements. The grey data point in the lower left of the graph represents another foetal/neonatal specimen that will not be discussed further. This sample is smaller than the others, but displays the now familiar pattern of adults and immatures (Group C), a single older juvenile in the middle (B), and three younger juveniles in the lower left (Group A). The two clusters of juvenile measurements indicate both early winter and spring hunting.
DISCUSSION
21 This method relies on individuals in their first year of life in order to establish season of death and thereby determine the season of past hunting activity. Sample sizes of measurable femora from the midden deposits discussed here range from eleven specimens at Feature 73 to 32 specimens at Feature 2. Juveniles form less than half of the population in each case. Seasonal interpretations are therefore based on relatively small samples of measurements. Nonetheless, the data outlined above show that this metrical approach has enough resolution to not only identify juveniles vs. immatures and adults, as Storå (1994) demonstrated using the modern harp seal skeletons from the NRM, but also to distinguish between juveniles in their first spring, and those in their first winter. Both young pups (roughly two to three months old) and older juveniles (roughly ten months old) are represented in all three middens. This provides the first conclusive evidence of early winter harp seal hunting at Phillip’s Garden.
22 Today, harp seals are found in the Port au Choix region during both of their migrations, but they appear less reliably and in smaller numbers in early winter than in the spring. This suggests that the early winter harp seal hunt at Phillip’s Garden may have been less predictable and less profitable than the spring one. At Feature 49, early winter juveniles outnumber spring juveniles four to three. In the other two features, on the other hand, spring juveniles outnumber early winter ones. At Feature 2, there are eight spring pups and only five winter ones, and at Feature 73, the spring kill is indicated by three specimens and the early winter kill by a single specimen. However, the small sample sizes prevent any firm conclusions about the relative importance of the two hunting seasons.
23 Harp seal dominates the faunal remains from Phillip’s Garden to such an extent that the Dorset diet at the site must have consisted almost exclusively of this species. The measurement data indicate that throughout the history of Phillip’s Garden, it was used for sealing both in the early winter and the spring. However, this does not mean that both hunts were necessarily pursued every year. The presence of an early winter seal hunt at Phillip’s Garden helps to expand our knowledge of Dorset settlement and subsistence practices on Newfoundland’s northwest coast. For the first time we can confidently place the Dorset at the outer coast in early winter. Whether this behaviour was consistent from year to year, and where they spent the winter months following the harp seal migration remain open to question. Perhaps they moved across the Great Northern Peninsula to the Island’s northeast coast. Schwarz (1994) has suggested that the occupants of Phillip’s Garden might have stored enough harp seal in December to carry them through to the following spring. Unfortunately, metrical analysis cannot resolve this issue since any stored seal meat consumed in March would still bear the metrical signature of a December kill. However, the specialized nature of the faunal assemblages and the lack of large pits, caches, or other significant storage features at the site suggest that the site was not frequently occupied during periods when the harp seal were unavailable. Thus, while the details and the full range of variability of the Dorset’s annual movements remain to be determined, this study suggests that Phillip’s Garden played a more extensive and/or varied role than was previously realized, acting as a seal hunting centre not only in spring, but also in early winter.
Acknowledgements
I am greatly indebted to Priscilla Renouf for allowing me to work on the bone from her excavations at Phillip’s Garden, and for commenting on an earlier draft of this paper. Thanks also to two anonymous reviewers for their constructive criticisms. Funding for the author’s excavation of Feature 73 was provided by the J.R. Smallwood Foundation for Newfoundland and Labrador Studies, the Institute of Social and Economic Research at Memorial University, and the Newfoundland and Labrador Department of Tourism and Culture. Additional funds were received through SSHRC and Parks Canada grants awarded to Priscilla Renouf. Kendra Wheatley assisted in sorting some of the measurable bone from the rest of the samples. This analysis was made possible by a post-doctoral fellowship from the Social Sciences and Humanities Research Council of Canada.References
Bowen, W.D., D.E. Sergeant, and T. Øritsland (1983), “Validation of Age Estimation in the Harp Seal, Phoca groenlandica, using dentinal annuli.” Canadian Journal of Fish-eries and Aquatic Science 40: 1430-1441.
Bull, John L. and J. Farrand (1994), National Audubon Society Field Guide to North American Birds. Eastern Region. New York: Knopf.
Harp, Elmer (1976), “Dorset Settlement Patterns in Newfoundland and Southeastern Hudson Bay.” In Eastern Arctic Prehistory: Paleoeskimo Problems, edited by Moreau M. Maxwell, pp. 119-138. Washington, DC: Memoirs of the Society for American Archaeology.
Hiseler, George, (1997), “Harp Seal (Phoca groenlandica) Canine Dentine Incremental Annuli as Indicators of Age And/Or Season of Death.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
Hodgetts, Lisa M. (1999), “Animal Bones and Human Society in the late Younger Stone Age of Arctic Norway.” PhD thesis, Department of Archaeology, University of Durham.
Hodgetts, Lisa M. (2001), “Seal Exploitation and Intra-Site Variation in the Younger Stone Age of Varangerfjord, North Norway.” In Archaeological Sciences ’97, edited by Andrew Millard, pp. 109-115. BAR International Series 939. Oxford: Oxbow Press.
Hodgetts, Lisa M. (2002), Report on the 2001 Excavations at Phillip’s Garden, Port au Choix National Historic Site. Ms. on file, Parks Canada, Halifax.
Hodgetts, L.M., M.A.P. Renouf, M.S. Murray, D. McCuaig-Balkwill, and L. Howse (2003), “Changing Subsistence Practices at the Dorset Palaeoeskimo Site of Phillip’s Garden, Newfoundland.” Arctic Anthropology 40 (1): 106-120.
LeBlanc, Sylvie (1996), “A Place With a View: Groswater Subsistence-Settlement Patterns in the Gulf of St. Lawrence.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
LeBlanc, Sylvie (1999), “Middle Dorset (1900-1100 BP) Regional Variability on the Island of Newfoundland and in Saint-Pierre et Miquelon.” In Identities and Cultural Contacts in the Arctic, edited by Martin Appelt, Joel Berglund, and Hans-Christian Gulløv, pp. 97-105. Copenhagen: Danish Polar Centre Publications.
LePage, A. (1989), “Une Transition Technique, les «Pêches» au Loup-Marin sur la Côte du
Labrador Depuis le Début du XVIII Siècle.” Anthropologie et Sociétés 13 (2): 55-78.
Maxwell, G. (1967), Seals of the World. London: Constable and Company Ltd.
Renouf, M.A.P. (1986a), Report of the 1985 Excavations at the Pointe Riche and Phillip’s Garden sites, Port au Choix National Historic Park. Ms. on file, Parks Canada, Halifax.
Renouf, M.A.P. (1986b), Port au Choix Resource Interviews. Unpublished field notes on file at the Archaeology Unit, Memorial University of Newfoundland.
Renouf, M.A.P. (1987), Archaeological Excavations at the Port au Choix National Historic Park. Report of the 1986 Field Activities. Ms. on file, Parks Canada, Halifax.
Renouf, M.A.P. (1991), Archaeological Investigations at the Port au Choix National Historic Park: Report of the 1990 Field Activities. Ms. on file, Parks Canada, Halifax.
Renouf, M.A.P. (1993), The 1992 Field Season, Port au Choix National Historic Park: Report of Archaeological Excavations. Ms. on file, Parks Canada, Halifax.
Renouf, M.A.P. (2000), “Symbolism and Subsistence: Seals and Caribou at Port au Choix, Northwestern Newfoundland.” In Animal Bones, Human Societies, edited by P.A. Rowley-Conwy, pp. 65-73. Oxford: Oxbow Books.
Renouf, M.A.P. (2002), Archaeology at Port au Choix, 1990 to 1992 Excavations. Occasional Papers in Northeastern Archaeology, 12. St. John’s: Copetown Press.
Renouf, M.A.P. and M.S. Murray (1999), “Two Winter Dwellings at Phillip’s Garden, A Dorset site in Northwestern Newfoundland.” Arctic Anthropology 36 (1-2): 118-132.
Robbins, Douglas T. (1985), “Stock Cove, Trinity Bay: The Dorset Eskimo Occupation of Newfoundland from a Southeastern Perspective.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
Schwarz, Frederick A. (1994), “Paleo-Eskimo and Recent Indian Subsistence and Settlement Patterns on the Island of Newfoundland.” Northeast Anthropology 47: 55-70.
Sergeant, D.E. (1991), Harp Seals, Man and Ice. Canadian Special Publication of Fisheries and Aquatic Sciences 114. Ottawa: Fisheries and Oceans Communications Directorate.
Storå, Jan (1994), “Åldersbedömning av sälben. En relativ fusioneringssekvens för grönlandssäl Pagophilus groenlandicus samt et analys av åldersstrukturen för benmaterialet från Jettböle stenåldersboplats på Åland.” Undergraduate diss., Osteological Research Laboratory, Stockholm University.
Storå, Jan (2002), “Neolithic Seal Exploitation on the Åland Islands in the Baltic Sea on the Basis of Epiphyseal Fusion Data and Metric Studies.” International Journal of Osteoarchaeology 12: 49-64.
Weber, A., O.I. Goriunova, and A.K. Konopatskii (1993), “Prehistoric seal hunting on Lake Baikal: methodology and preliminary results of the analysis of canine sections.” Journal of Archaeological Science 20: 629-644.
Notes