Animal Exploitation and Season of Occupation at the Groswater Palaeoeskimo Site of Phillip’s Garden West
Patricia J. WellsArchaeology Unit of Memorial University
INTRODUCTION
1 ANIMAL BONES from archaeological sites can contribute to an understanding of past cultures. Determining what species were exploited and during what season(s) enables a discussion of the settlement and subsistence practices of prehistoric populations. This paper examines faunal remains from the Groswater Palaeoeskimo site of Phillip’s Garden West (EeBi-11) at Port au Choix (Figure 1). Its aim is to understand the nature of site occupation and hunting activities that took place there. While authors have suggested models for Groswater Palaeoeskimo site placement and the animals likely exploited (Fitzhugh 1972; Loring and Cox 1986), there are only a few sites from which faunal remains have been recovered and analyzed, and which can therefore test these models. Evidence from faunal remains collected at Phillip’s Garden East (EeBi-1) and Factory Cove (DlBk-3) further south on the Great Northern Peninsula (Figure 1) show a strong focus on seal hunting supplemented by small amounts of fish, birds and terrestrial mammals (Auger 1985; Kennett 1991).
2 Phillip’s Garden West is unique in the Groswater archaeological record because a number of the stone tool types found here morphologically differ from tools typical of this culture (Ryan 1997; Renouf 2005). For instance, endblades and bifaces are often much longer and thinner than typical examples, and display unusually fine edge serration and surface grinding (Renouf 2005). One tentative explanation for the difference is that this site was occupied during a different season from other Groswater sites in the region (Renouf 1994), thereby necessitating a different kind of tool kit. Identification of the faunal remains from this site will establish the species exploited and at what time of year. In addition, a review of the ecology of these species will allow a discussion of Groswater hunting practices.
Figure 1. Location of sites mentioned in the text.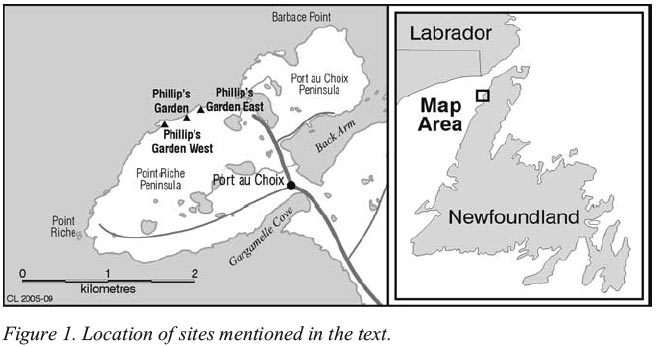
PHILLIP’S GARDEN WEST
3 Phillip’s Garden West is on the Point Riche Peninsula situated less than half a kilometre west of a second Groswater Palaeoeskimo site, Phillip’s Garden East (Figure1). Phillip’s Garden West was tested in 1984 and completely excavated during the summers of 1990-1992 (Renouf 1985, 1991, 1992, 1993). The site is on a 13-metre terrace and, with an area of about 500 square metres, almost covers the terrace slope. A small living floor, interpreted as a tent structure, was outlined by five post-holes. One hearth was inside the structure and four more outside. Charcoal samples from the site range from 2680 to 1820 cal BP.1
4 Below the terrace edge the land slopes steeply to the beach, and is covered by refuse thrown from the upper terrace. The bulk of the artifactual and faunal material was recovered from these hillside midden deposits. Despite the mixing that would have taken place as refuse was thrown over the hillside, the excavators were able to distinguish separate dumping episodes.
5 Renouf (1992, 1998) suggested that the site might represent a warm weather occupation as the living floor is located on an exposed terrace and most of the hearths were outdoor fires. Furthermore, there are few fire-cracked rocks — those rocks that bear the signs of heating, such as cracks and reddening — which were ubiquitous at Phillip’s Garden East which was shown to have been occupied during the cold weather months (Renouf 1994). Also, there were no soapstone lamps found on this site which, although rare on Groswater sites, were found at Phillip’s Garden East, and would have been used for heat and light. The faunal evidence from Phillip’s Garden West directly addresses the question of season of occupation.
ENVIRONMENT AND RESOURCES
6 The Groswater Palaeoeskimos of Port au Choix lived in an environment that was colder and less predictable than today (Rosenberg et al. this volume). Environmental conditions would have influenced the availability and frequency of animal resources in this region. While it is possible that there were differences in the past, it is assumed that the range of species available in Newfoundland today is much the same as it was in the past, and that the ecology of these species has not changed significantly.
7 Available animal resources would have included species present today. Species that are rare today, such as the ringed seal (Phoca hispida) and walrus (Odobenus rosmarus), could have been more frequent in the past. A wide variety of birds would have been available for exploitation. In addition to huge colonies of nesting murres (Uria sp.), auks (Pinguinus impennis), and razorbills (Alca torda) found on offshore islands, a range of freshwater and salt-water ducks, geese, gulls, loons (Gavia sp.), and ptarmigan (Lagopus sp.) would have been present. Marine and freshwater fish would have been abundant, while sea mammals would have provided large quantities of meat, fat, and hide as well as bone and ivory. Large terrestrial species are few in Newfoundland, but one, the caribou (Rangifer tarandus), would have been of great value for its meat, grease, marrow, hide, and antler. Fur-bearing mammals would have been available, providing hide as well as meat.
8 The flora of Newfoundland would have provided a wide variety of berries (see Deal this volume). While these would surely have made up part of the Groswater Palaeoeskimo diet, lack of preservation in the archaeological record does not allow a full appreciation of the role of this foodstuff. Nonetheless, animal protein almost certainly made up the bulk of the diet of the Groswater people.
THE FAUNAL SAMPLES
9 Three faunal samples were selected from well-defined and dated midden features that span the occupation of the site. Feature 18 is the earliest midden sample with two radiocarbon dates, 2700-2360 cal BP (Beta 49761)2 and 2680-2160 cal BP (Beta 49760). Approximately half of the material in this large feature was analyzed. All of features 5A-5D and 5E were examined. They are dated to 2330-2150 cal BP (Beta 66437) and 2000-1820 cal BP (Beta 66438), respectively. Together the samples represent approximately two-thirds of the total faunal remains from the site.
10 The first stage in understanding the settlement and subsistence patterns of the Groswater Palaeoeskimo at Phillip’s Garden West is to document the species exploited. This involves identifying the species present and establishing their relative abundance. The species that are available in the region during a restricted time of the year helps establish the season(s) of occupation both at one point in time and over time.
PRESERVATION
11 Faunal preservation at Phillip’s Garden West is excellent, partially due to the limestone bedrock which neutralizes the otherwise acidic soil common in Newfoundland. Nonetheless, it is possible that preservation conditions may not be the same for each of the individual faunal assemblages; thus, observed differences in species abundance could be the result of natural destruction, rather than reflect differences in subsistence activity at the site over time. Particularly with the tiny bones of fish and birds, slight differences in preservation conditions can influence the relative frequency of these species. One way to recognize differences in preservation is to calculate the ratio of identified to unidentified fragments of bone for each of the bone assemblages (Hodgetts et al. 2003). Because humans and carnivores often contribute to the fragmentation of bone, breaking it up for marrow and grease, this technique is not always appropriate. However, there was no evidence of carnivore activity at Phillip’s Garden West. Furthermore, since seal bones dominate these assemblages and their dense structure means they have little marrow and grease, natural fragmentation is the most likely destructive agent (Hodgetts et al. 2003). Table 1 shows that the assemblages have similar ratios of identified to unidentified fragments. The variation in species abundance for each assemblage is thus unlikely to be a reflection of differential preservation.
Table 1. Identified to unidentified ratios of fragments for faunal assemblages at Philip's Garden West

METHODOLOGY
12 The Phillip’s Garden West faunal material was identified using comparative collections of animal skeletal material at the Provincial Museum of Newfoundland and Labrador and the Canadian Museum of Nature. The first level of identification was the bone (element), as well as its side (right or left) in appropriate cases; the second was to determine the most precise taxonomic level possible for each element. Often it is difficult to differentiate species for every element in the body. For instance, with few exceptions, there is a considerable overlap in the appearance of elements among seal species (Hodgetts 1999). Thus, my identifications of phocids were often limited to the family level, with some narrowed down to two species, and only a few identified to a single species. There were many tiny fragments of bone in each of the samples that could only be given a class designation (i.e., mammal, fish, bird).
13 Establishing the relative abundance of different species on a site involves counting the fragments and whole elements. The most basic method of counting bones is to identify the species from which the bone came and to count the total number of bones for that species. The number generated is the Number of Identifiable Specimens (NISP), with a specimen being defined as a whole or portion of a skeletal element (Reitz and Wing 1999: 10). NISP counts are a descriptive means of organizing and presenting a bone assemblage.
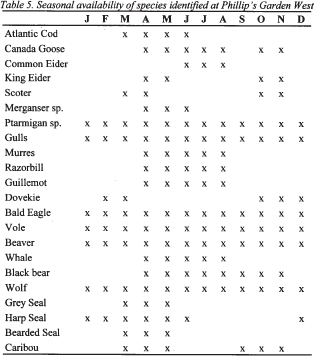
14 When trying to understand how many individual animals of each species are represented by the bones in an assemblage, a calculation, the Minimum Number of Individuals (MNI), gives the minimum number of animals which can be accounted for in a collection. To calculate the MNI, bones for each taxon are grouped according to element and divided into the side of the animal from which they came. The highest number represents the MNI. Considering the age, sex, and size of the bones can influence the MNI count since it increases the basis for differentiation. For example, if humeri (upper front limb bones) were the most frequent bone for a taxa, with ten left and four right elements, the MNI based on side would be ten. This number changes when age is considered. If the four right humeri were unfused (growth unfinished, therefore representing younger individuals) and the ten left fused (representing older individuals), they could not have come from the same individual and the MNI for this taxa would become fourteen.
15 The fragmented state of many specimens in the archaeological record can inflate the NISP and MNI. When an element is fragmented, the same bone can be counted a number of times. One way to overcome this problem is to give recognizable parts on each element zonal designations, and when identifying bones, record the zones that are present. The most frequently represented zone will give the minimum number of that particular element.
16 The calculation of MNI in this paper takes side and fusion into consideration. In addition, element size was considered for the fish bones only. For each taxonomic designation, the most frequent zone of the most frequent element generated the MNI for that taxon. Once the side of the animal from which this element came was taken into consideration, differences in fusion states for that element allowed for the possibility of increasing the MNI.
SPECIES ABUNDANCE
17 Tables 2, 3, and 4 present the species abundance for each of the three feature assemblages in this study. Faunal assemblages from the midden features at Phillip’s Garden West contain a range of species dominated by sea mammals, especially seal, with a variety of birds, some terrestrial mammals and fish. The most numerous bird species are those of the Family Alcidae (sea birds) including dovekie (Alle alle), common and thick-billed murre (Uria aagle, Uria lomvia), razorbill (Alca torda), and guillemot (Cephus grylle).
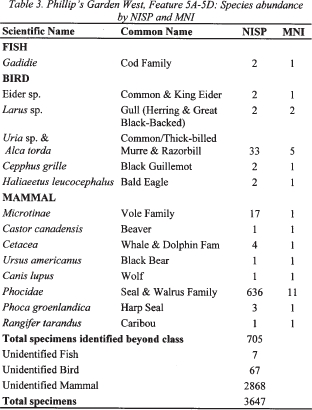
18 Seal species dominate the faunal assemblage from all features at Phillip’s Garden West. A number of seal species are available in the area; however, it is the predictable migration of huge numbers of harp seal (Phoca groenlandicus) during the spring and winter that probably provided the bulk of the seal meat at this site. Harp seals dominate among those Phocidae bones that could be identified to species. Grey (Halichoerus grypus) and bearded seals (Erignathus barbatus) were also available, but mostly as isolated individuals which were probably exploited in an opportunistic way. Harp seals were the main focus at this site. Other sea mammals identified include a few specimens from the family Cetacea (whales).
19 A number of terrestrial mammals were found in the midden features. The smallest of these was the vole, which is likely to have died of natural causes. While only a few caribou specimens were identified, a number of the unidentifiable bone fragments had the texture of terrestrial rather than marine mammal bones, suggesting that some fragments are likely to be caribou. Caribou long bones were often processed for the extraction of marrow and bone grease and therefore end up as fragments (Lyman 1991; Hodgetts 1999). This in turn suggests that there may have been more caribou in the diet at this site than is apparent from the remains. The presence of the identifiable elements, as well as the apparent shattered terrestrial mammal long bones, suggests that some portions of caribou could have been transported to this site, providing nourishment until the arrival of the harp seal herds. The other terrestrial mammals found in the assemblage were red fox (Vulpes vulpes), wolf (Canis lupus), black bear (Ursus americanus) and beaver (Castor canadensis).
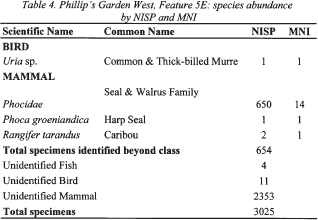
20 The only fish identified from the midden features was the Atlantic cod (Gadus morhua). Most of the identifications were made on vertebrae, and, based on their size, a MNI of six for Feature 18 and one for Feature 5A-5D was generated.
21 By examining faunal samples from different time periods it is possible to compare subsistence activities over time, including the differences in species diversity. While it might appear that during the earliest occupation of Phillip’s Garden West (Feature 18) the Groswater Palaoeskimos were exploiting a greater variety of species than later in the occupation (Features 5A-5D, 5E), in fact this is not the case. The samples from Feature 5A-5D and Feature 5E are much smaller than Feature 18 which means that less common taxa are unlikely to be represented. Samples need to consist of at least 1,000 identifiable specimens in order to represent the entire species range with confidence (Grayson 1984). While it is possible that the species variation in these three assemblages could be the result of real change over time, most likely sample size accounts for the differences in species diversity. In fact the results suggest that at Phillip’s Garden West there is not a great deal of variation in the nature of subsistence over time.
Table 2. Philip's Garden West, Feature 18: Species Abundance by NISP and MNI

Display large image of Table 2
Table 3. Philip's Garden West, Feature 5A-5D: Species abundance by NISP and MNI
Display large image of Table 3
Table 4. Philip's Garden West, Feature 5E: species abundance by NISP and MNI
SEASONALITY
22 When the results of these identifications are compared to modern species ecology, a spring-early summer and late fall-early winter occupation of Phillip’s Garden West is suggested. Table 5 shows the seasonal availability of the species present.
23 Many of the bird species were available during both spring and fall, indicating site occupation during these periods. The exception is the dovekie, which is common during the fall and found only occasionally early in the spring in ice-free channels along the shore (Nettleship and Birkhead 1985). In the spring it would be possible to harvest both eggs and birds in the Port au Choix region, while in the early summer young birds could be preyed upon. Indeed, one of the razorbill elements identified was from a young individual, strengthening the suggestion of a spring-early summer hunt. Murres and razorbills are common in onshore waters during the spring and summer, and become much less common into the summer as they move offshore (Nettleship and Birkhead 1985). The king eider (Somateria spectabilis) is available during the spring and fall, while the common eider (Somateria mollisima) is strictly a summer visitor to the region (Godfrey 1966: 75). Common and red-breasted mergansers (Mergus sp.) are available during the spring only, while the scoter species (Melanitta sp.) are present during the spring and fall (Threlfall 1983). The species available year-round include gulls, rock and willow ptarmigan, and the bald eagle (Haliaeetus leucocephalus). The gull elements could only be identified to the genus level, but they were all large, suggesting great black-backed (Larus marinus), herring (L. argentatus) or possibly glaucous gulls (L. Hyperboreus). As gulls are not useful seasonal indicators, this lack of precision does not affect the interpretation of seasonality. The ptarmigan is the only truly terrestrial bird species identified. The willow (Lagopus lagopus) and rock ptarmigan (Lagopus mutus) are present year-round; however, the rock ptarmigan occurs mainly on higher ground on the Long Range Plateau (Godfrey 1966; John Wells pers. comm. 2001).
Table 5. Seasonal availability of species identified at Philip's Garden West
Display large image of Table 5
24 Harp seals spend their summers in the arctic. In fall, a large proportion of them migrates to the Gulf of St. Lawrence just ahead of winter ice formation (Sergeant 1991). The Gulf herd passes through the Strait of Belle Isle in winter during December or January. Birth, breeding, and molting take place in early spring on the pack ice in the Gulf of St. Lawrence and occasionally, when conditions create firm ice, off Mecatina Island on the Quebec Lower North Shore (Sergeant 1991: 41; Stenson et al. 1995). Northward migration of the harp seals takes place from late April until early June (Sergeant 1991: 85). During this season, harp seals feed on capelin and cod found near the coast (LeBlanc 1996).
25 It is likely that harp seals were hunted from Phillip’s Garden West during both the early winter and spring. Fourteen foetal or neonatal seal elements were identified in Feature 18. This suggests possible hunting of pregnant females in the late fall-early winter as the seals made their way south. However, these numbers are small, and because it is extremely difficult to distinguish foetal from neonatal elements, it cannot be ruled out that these bones represent newborns heading north in the early spring. Distinguishing a spring from an early winter hunt is difficult.
26 Hodgetts (this volume) uses limb bone measurements to determine the seasonality of harp seal hunting at the Dorset site of Phillip’s Garden. Since harp seals are born during the spring only, a population will have a particular configuration of age groups, and therefore limb sizes, at any given time. A spring herd will include 2-3-month juveniles, immature individuals and adults, the latter two groups indistinguishable through bone measurement and thus forming a single size-age class. A December herd will also consist of two age classes: 10-month juveniles and immature adults. In a number of faunal assemblages from Phillip’s Garden, Hodgetts identified three age groups: newborns, juveniles, and immature adults, thus demonstrating seal hunting during both times of the year. While these measurements have not yet been done for the Groswater samples from Phillip’s Garden West, it is reasonable to suggest that the seasonal hunting pattern was similar for Groswater Palaeoeskimos.
27 Although terrestrial mammals make up a small portion of the Phillip’s Garden West assemblages, some seasonal information is suggested. The caribou could have been exploited during the winter when they move over the barrens, or in spring during their migration toward calving grounds in the interior (Cameron 1958; Bergerud 1983; Earl Pilgrim, pers. comm. 2001). The black bear, wolf, fox, and beaver would have been most valuable during the winter when their furs are at their best. However, they would have been available year-round in the general area. There may have been less forest cover during the Groswater Palaeoeskimo occupation of the area. This would not affect the availability of wolf, black bear, or fox to any great extent, as these animals are found in a range of habitats (Cameron 1958; Northcott 1974; Forsyth 1985). The beaver would have been rare in the region because of its dependence on forests for food and shelter. Tooth fragments only represent the black bear and beaver. It is possible that these teeth were used as tools for hide working, or incising jobs, or possibly as amulets (Tyzzer 1943). Consequently, it is possible that the black bear and beaver were hunted elsewhere and only their teeth transported to the site for uses other than subsistence. The wolf, black bear, red fox, and beaver do not provide conclusive seasonal evidence.
28 All the cod vertebrae identified are from large individuals of over 60 centimetres in length (~ 2.7 kg). Cod are most commonly found in deeper waters; however, they will follow capelin (Mallotus villosus) and shrimp (Pandalus borealis) close to shore in the spring, rising to shallower depths until the summer heat drives them offshore into deep water. It is most likely that cod was exploited in shallow water in the spring.
29 The faunal remains from Phillip’s Garden West suggest an occupation during the spring-early summer and late fall-early winter. This is based on modern animal ecology of the species identified at the site. The arrival each spring and early winter of large harp seal populations probably directed the seasonal arrival of the Groswater Palaeoeskimos to the region. The other migratory species have similar seasonal migratory patterns as the harp seal, supporting occupation during these periods. Because the dominant focus on seal did not change over time, it is likely that seasonality was the same throughout the occupation of the site.
HUNTING AT PHILLIP’S GARDEN WEST
30 Hunting at Phillip’s Garden West is suggested based on the ecology of the species exploited and the known material culture of the Groswater Palaeoeskimo, in particular harpoon technology. Many of the species exploited are migratory and available for restricted periods at relatively predictable times during the year. It is likely that the Groswater people moved into this region in anticipation of the arrival of these species.
31 Evidence for Groswater Palaeoeskimo fishing is sparse. Little is known about the technology employed in the capture of cod. No fishhooks or other evidence of fishing technology has ever been recovered from Groswater Palaeoeskimo sites.
32 In a now famous letter written in 1497, John Cabot described waters off the coast of Newfoundland as “swarming with fish, which can be taken not only with the net, but in baskets let down with a stone” (Pope 1997: 27). There is little ethnographic evidence for fishing nets being used by Inuit hunters before European trade (Mathiassen 1927: 58). Nevertheless, it is possible that the Groswater Palaeoeskimos had a technology such as baskets or, more likely, nets for capturing cod. While nets would not survive in the archaeological record, the presence of two stone net-sinkers at the nearby Groswater site of Phillip’s Garden East strengthens the suggestion that nets were used (Renouf 1998). If fish hooks had been used, it is presumed that they would have been made from a strong material such as bone or ivory. Their absence from the archaeological record at Phillip’s Garden West, where faunal preservation is excellent, suggests they were not manufactured or deposited at this site. There were no barbed bone points found in the Phillip’s Garden West tool assemblage to indicate leister (fish spear) use, but this is an unlikely method of cod fishing as cod rarely come close to the surface, preferring depths of approximately 5 metres in inshore regions (Lear 1989).
33 The majority of bird species exploited at Phillip’s Garden West are seabirds found offshore on islands in the region. It is likely that they were exploited from boats or on foot. It is not clear how these species would have been killed. Groswater material culture does not include clearly discernible bird hunting technology; however, it is possible that this technology is present but not recognized.
34 Terrestrial mammal hunting was not a major focus at Phillip’s Garden West. Caribou may have been of greater importance at other times of the year and in other locations. Although many of the fragments of unidentifiable mammal from Phillip’s Garden West could have been caribou, these fragments are not so numerous as to suggest an intentional hunt to return large quantities of caribou to this site.
35 It seems likely that Groswater Palaeoeskimo hunters used boats to hunt seals along the edge of the pack ice using harpoons fitted with stone endblades. Ice conditions in these areas today are usually too trecherous to risk travel on foot (LeBlanc 1996); however, fishermen from the area are known to have walked out onto the ice to hunt harp seals in the spring of 2003. In the past, conditions may have allowed hunters access to seals on the ice on a more consistent basis. In addition, it is possible that large groups of seals hauled out on the beaches around the Port au Choix and Point Riche peninsulas. While harp seals seldom haul out on land, Port au Choix resident John Gould noted that numerous harp seals were seen on local beaches, including the beach at Phillip’s Garden, in recent years. Earl Pilgrim, a resident of Roddickton on the east coast of the Great Northern Peninsula, reported numerous harp seals landing on beaches around the community in winter. Therefore, it seems possible that in the past, hunting seals from the land could have been accomplished by individual hunters or by groups of hunters, on foot.
36 A small number of whale (family Cetacea) elements were identified in the feature assemblages (NISP 5 for all features). These morphologically distinct elements are readily identifiable in faunal assemblages, and their relative scarcity suggests that whale exploitation was minimal. Indeed it may only have involved opportunistic scavenging. All of the whale elements were from small whales. The sample of whale elements is too small to make convincing statements about methods of retrieval; nevertheless, if scavenging were the likely method of obtaining whalebone, one would expect the presence of whale elements of varying sizes. The consistent small size of the whale elements from the Phillip’s Garden West assemblages suggests that there could have been occasional opportunistic hunting of small whales using seal hunting technology. While this argument is compelling, it remains speculative.
CONCLUSION
37 The faunal remains from Phillip’s Garden West contribute to an interpretation of Groswater Palaeoeskimo settlement and subsistence at this site. The analysis demonstrates the importance of seal, particularly harp seal, to the inhabitants’ diet, a focus that did not change over the period of site occupation. Other species that were available at approximately the same time of year supplemented the diet to a lesser extent. The availability of harp seal in the region dictated the seasons of site occupation. In modern times they are available twice during the year for restricted periods as they migrate to the Gulf of St. Lawrence from the Arctic. The data suggest that Groswater Palaeoeskimo people occupied Phillip’s Garden West during the spring and winter.
38 It was earlier proposed (Renouf 1994) that the unique morphology of the Phillip’s Garden West stone tools might be the result of a warm weather occupation rather the winter and spring occupation of the nearby Groswater site, Phillip’s Garden East. This study shows that both sites were occupied during the same time of year and therefore other interpretations of the Phillip’s Garden West tools must be sought (Wells 2002; Renouf 2005).
Acknowledgements
Funding for this research was provided by the Smallwood Foundation for Newfoundland and Labrador Studies and the Institute of Social and Economic Research. Priscilla Renouf and Lisa Hodgetts gave valuable assistance in the preparation of the research. Assistance with some identification was provided by Darlene Balkwill and Steven Cumbaa, both of the Canadian Museum of Nature in Quebec. I extend thanks to two anonymous reviewers for their helpful comments.References
Auger, R. (1985), “Factory Cove: Recognition and Definition of the Early Palaeo-Eskimo Period in Newfoundland.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
Bergerud, A.T. (1983), “Prey Switching in a Simple Ecosystem.” Scientific American 249(6): 130-141.
Cameron, A.W. (1958), Mammals of the Islands in the Gulf of St. Lawrence. National Museum of Canada Bulletin 154, Ottawa.
Deal, M. (this volume), “Palaeoethnobotanical Research at Port au Choix.”
Fitzhugh, W.W. (1972), Environmental Archaeology and Cultural Systems in Hamilton Inlet, Labrador. Smithsonian Contributions to Anthropology 16, Washington, DC.
Forsyth, A. (1985), Mammals of the Canadian Wild. Camden East: Camden House Publishing Ltd.
Godfrey, W.E. (1966), The Birds of Canada. National Museum of Canada Bulletin No. 203, Ottawa.
Grayson, D.K. (1984), Quantitative Zooarchaeology: Topics in the Analysis of Archaeological Faunas. New York: Academic Press.
Hodgetts, L.M. (1999), “Animal Bones and Human Society in the Late Younger Stone Age of Arctic Norway.” PhD thesis, Department of Archaeology, University of Durham.
Hodgetts, L.M. (this volume), “Using Bone Measurements to Determine the Season of Harp Seal Hunting at the Dorset Palaeoeskimo Site of Phillip’s Garden.”
Hodgetts, L.M., M.A.P. Renouf, M.S. Murray, D. McCuaig-Balkwill, and L. Howse (2003), “Changing Subsistence Practices at the Dorset Palaeoeskimo Site of Phillip’s Garden, Newfoundland.” Arctic Anthropology 40 (1): 106-120.
Kennett, B.L. (1991), “Phillip’s Garden East: An Examination of The Groswater Palaeo-Eskimo Phase.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
LeBlanc, S. (1996), “A Place with a View; Groswater Subsistence-Settlement Patterns in the Gulf of St. Lawrence.” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
Lear, W.H. (1989), Underwater World. Atlantic Cod. Communication Directorate Department of Fisheries and Oceans, Ottawa.
Loring, S. and S. Cox (1986), “The Postville Pentecostal Groswater Site, Kaipokok Bay, Labrador.” In Palaeo-Eskimo Cultures in Newfoundland, Labrador, and Ungava, pp. 65-94. Memorial University of Newfoundland Reports in Archaeology 1, St. John’s.
Lyman, R.L. (1991), “Taphonomic Problems with Archaeological Analysis of Animal Carcass Utilization and Transport.” In Beamers, Bobwhites, and Blue-points: Tributes to the Career of Paul W. Parmalee, edited by J.R Purdue, W.E. Klippel, and B.W. Styles, pp.125-138. Illinois State Museum Scientific Papers, Vol. 23. Springfield.
Mathiassen, T. (1927), The Thule Culture and its Position Within the Eskimo Culture: Archaeology of the Central Eskimo II. Report of the Fifth Thule Expedition 1921-24. Copenhagen: Gyldendalske Boghandel.
Nettleship, D.N. and T.R. Birkhead (1985), The Atlantic Alcidae: The Evolution, Distribution, and Biology of the Auks Inhabiting the Atlantic Ocean and Adjacent Water Areas. London: Academic Press.
Northcott, T.H. (1974), The Land Mammals of Insular Newfoundland. Wildlife Division, Department of Tourism, Culture and Recreation, Government of Newfoundland and Labrador, St. John’s.
Pope, P.E. (1997), The Many Landfalls of John Cabot. Toronto: University of Toronto Press.
Reitz, E.J. and E.S. Wing (1999), Zooarchaeology. Cambridge Manuals in Archaeology. Cambridge: Cambridge University Press.
Renouf, M.A.P. (1985), Archaeology of the Port au Choix National Historic Park: Report of the 1984 Field Activities. Ms. on file, Parks Canada, Halifax.
Renouf, M.A.P. (1991), Archaeological Investigations at the Port au Choix National Historic Park: Report of the 1990 Field Activities. Ms. on file, Parks Canada, Halifax.
Renouf, M.A.P. (1992), The 1991 Field Season, Port au Choix National Historic Park: Report of Archaeological Excavations. Ms. on file, Parks Canada, Halifax.
Renouf, M.A.P. (1993), The 1992 Field Season, Port au Choix National Historic Park: Report of Archaeological Excavations. Ms. on file, Parks Canada, Halifax.
Renouf, M.A.P. (1994), “Two Transitional Sites at Port au Choix, Northwestern Newfoundland.” In Threads of Arctic Prehistory: Papers in Honour of William E. Taylor Jr.,edited by D. Morrison and J-L. Pilon, pp. 166-195. Canadian Museum of Civilization, Mercury Series Paper 149, Hull.
Renouf, M.A.P. (1998), “Archaeology at Port au Choix Northwestern Newfoundland. Volume 2: 1990-1992.” Occasional Papers in Northeastern Archaeology 10: 3-47.
Renouf, M.A.P. (2005), “Phillip’s Garden West: A Newfoundland Groswater Variant.” In Contributions to the Study of the Dorset Palaeo-Eskimos, edited by P. Sutherland, pp. 57-80. Archaeological Survey of Canada Mercury Series, CMC Publications, Hull.
Rosenberg, S.M., I.R. Walker, and J.B. Macpherson (this volume), “Environmental Changes at Port au Choix as Reconstructed from Fossil Midges.”
Ryan, K. (1997), “Groswater Palaeo-Eskimo Tool-makers: Phillip’s Garden West and Beyond.” Hons. thesis, Department of Anthropology, Memorial University of Newfoundland.
Sergeant, D.E. (1991), Harp Seals, Man and Ice. Ottawa: Department of Fisheries andOceans.
Stenson, G.B., M.O. Hammill, M.C.S. Kingsley, B. Sjare, W.G. Warren, and R.A. Myers (1995), Pup Production of Harp Seals, Phoca groenlandica, in the Northwest Atlantic During 1994. Department of Fisheries and Oceans, Atlantic Fisheries Research Document 95/20, St. John’s.
Threlfall, W. (1983), “Seabirds.” In Biogeography and Ecology of the Island of Newfoundland, edited by G.R. South, pp. 467-508. Boston: W. Junk Publishers.
Tyzzer, E.E. (1943), “Animal Tooth Implements From Shell Heaps of Maine.” AmericanAntiquity 8: 354-362.
Wells P.J. (2002), “An Analysis of Faunal Remains from Two Groswater Palaeoeskimo Sites at Port au Choix, Northwestern Newfoundland: Phillip’s Garden West (EeBi-11) and Phillip’s Garden East (EeBi-1).” MA thesis, Department of Anthropology, Memorial University of Newfoundland.
Notes