Testing a Proposed Late Holocene Sea-Level Oscillation using the Isolation Basin Approach, Great Northern Peninsula, Newfoundland
I. Rod SmithResearch Scientist with Natural Resources Canada, Geological Survey of Canada, Calgary
Trevor Bell
Associate Professor of Geography at Memorial University
M. A.P. Renouf
Department of Geography, Memorial University
INTRODUCTION
1 THIS STUDY TESTS Grant’s hypothesis (1994) that northwestern Newfoundland experienced a 10-metre sea-level oscillation during the late Holocene,1 against previous reconstructions that propose continuous emergence. This study also comprises part of a larger regional project that is reconstructing spatial and temporal patterns of postglacial sea-level history for Newfoundland and Labrador (e.g., Bell et al. 2001 this volume) and assessing its archaeological implications (e.g., Bell and Renouf 2003; Smith et al. 2003). The importance of palaeoshoreline position for understanding prehistoric coastal settlement is well established in areas of dynamic sea-level change (e.g., Labrador, Clark and Fitzhugh 1992; Newfoundland, Bell and Renouf 2003; Gulf of Maine, Sanger and Belknap 1987, Sanger and Kellogg 1989; British Columbia, Josenhans et al. 1997). Studies show that reconstruction of palaeoshoreline elevation and configuration is an essential tool for explaining the location of known archaeological sites and for developing search strategies for finding others (Bell and Renouf 2003; Renouf and Bell in press).
2 To our knowledge, this is the first study in Newfoundland and Labrador that employs diatoms and lake basin isolation history to address questions related to sea-level change. Similar studies, however, have been widely used elsewhere in the world, including Scandinavia (Corner et al. 1999; Eronen et al. 2001), Greenland (Long et al. 1999), and western Canada (Hutchinson et al. 2004). The methodology involves identifying litho- and bio-stratigraphic indicators that accord with changes between marine and lacustrine sediments, in basins that have been isolated (or transgressed) by changing sea levels. This study uses diatoms, forms of aquatic algae that are particularly sensitive to changes in salinity. Once marine-lacustrine boundaries have been identified, they are dated using the radiocarbon method. This allows researchers to plot the elevation of outlets of various basins (thresholds for isolation/transgression) against time, and thus reconstruct relative sea-level fluctuations.
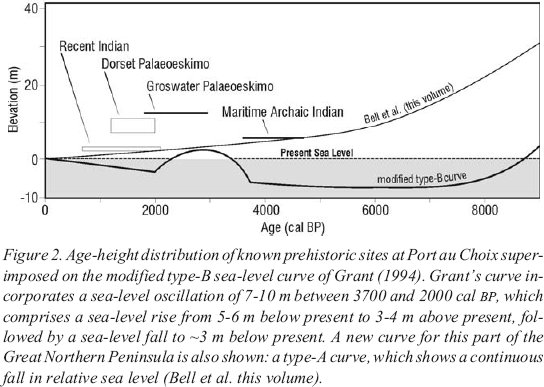
3 Our current understanding of RSL history for the Port au Choix region (Figure1) is presented in a companion paper in this volume (Bell et al. this volume), while the spatial pattern of sea-level change along the west coast of the Great Northern Peninsula is under review (Bell unpublished). These studies propose that Port au Choix is located near the transition between a continuously falling postglacial sea-level trend (type-A, Quinlan and Beaumont 1981) and one in which the postglacial sea level initially falls below its present level and then rises again (type-B, Quinlan and Beaumont 1981; Figure 2). Grant (1994) proposed a third possible sea-level history for the region, a modification of the type-B model, in which postglacial sea level falls below its present level at approximately 9000 cal BP,2 followed by a brief oscillation above present (reaching 3-4 m above sea level [asl]) between 3700 and 2000 cal BP, and a transgression to present sea level during the last several thousand years (Figure 2).
4 This theory was based on the occurrence of a low, horizontal relic sea cliff, which lies just above present tide level along the west coast of the Great Northern Peninsula (Figure 3; Grant 1994). The cliff is an erosional feature and so was not dated directly, but its age is apparently bracketed stratigraphically by 4500-year-old beaches at Port au Choix that it cuts into, and by 900-year-old beaches farther north that overlap it. Similar oscillations in sea level along the northeastern seaboard of North America have been attributed to the migration of a collapsing glacioisostatic forebulge (Barnhardt et al. 1995), eustatic changes related to mid-Holocene warming (Scott and Collins 1986) or, as Mörner (1976, 1992) has argued, to changes in the Earth’s geoid or rate of rotation.
5 The implications of Grant’s (1994) sea-level oscillation for the Port au Choix regional archaeological record were intriguing (Bell and Renouf 1998). Prior to recent finds, the only Maritime Archaic Indian (MAI) site at Port au Choix was a cemetery, radiocarbon dated between 4690 and 3620 cal BP (Tuck 1976) at 6-10 metres above sea level. The search for a habitation site associated with the cemetery was the focus of many unsuccessful surveys in the 1960s, 1980s, and 1990s. These site surveys were predicated on two assumptions. First, that camps or longer-term settlements would likely have been situated near the contemporaneous shoreline, and therefore at an elevation of 6-8 metres above present sea level, similar to the cemetery; and second, that the relative sea-level history of Port au Choix followed a simple type-A model. The proposition of another possible sea-level history for the region presented alternative search strategies for locating the MAI habitation site. For example, Grant’s (1994) modified type-B sea-level curve suggests that the Groswater and Dorset Palaeoeskimo sites would have been located 9-10 metres above the shoreline at the time of occupation, whereas the MAI cemetery, at 14-15 metres above sea level, would have been even farther removed from the active beach during its time of use (Figure 2). In other words, the cemetery would have been on higher ground than the Palaeoeskimo settlements. If MAI and Palaeoeskimo settlements were similarly situated from the contemporaneous shoreline, for similar practical considerations such as view, shelter and access, then MAI settlements might have been closer to the active beach than the cemetery. If MAI settlements were situated within 10 metres of the contemporary shore, as were the Palaeoeskimo sites, these MAI sites today would be at, or below, present sea level. Therefore the need for an accurate sea-level history to develop a search strategy for MAI habitation sites highlighted the importance of testing Grant’s proposed sea-level oscillation. Previous studies elsewhere had demonstrated the potential of the lake isolation method in resolving sea-level fluctuations of this magnitude, and so we adopted the approach as one part of an integrated geo-archaeological initiative in the Port au Choix region (Renouf and Bell 2002).
Figure 1. Location map for Gilmores and Otter ponds on the Great Northern Peninsula, northwest Newfoundland.Figure 2. Age-height distribution of known prehistoric sites at Port au Choix superimposed on the modified type-B sea-level curve of Grant (1994). Grant’s curve incorporates a sea-level oscillation of 7-10 m between 3700 and 2000 cal BP, which comprises a sea-level rise from 5-6 m below present to 3-4 m above present, followed by a sea-level fall to ~3 m below present. A new curve for this part of the Great Northern Peninsula is also shown: a type-A curve, which shows a continuous fall in relative sea level (Bell et al. this volume).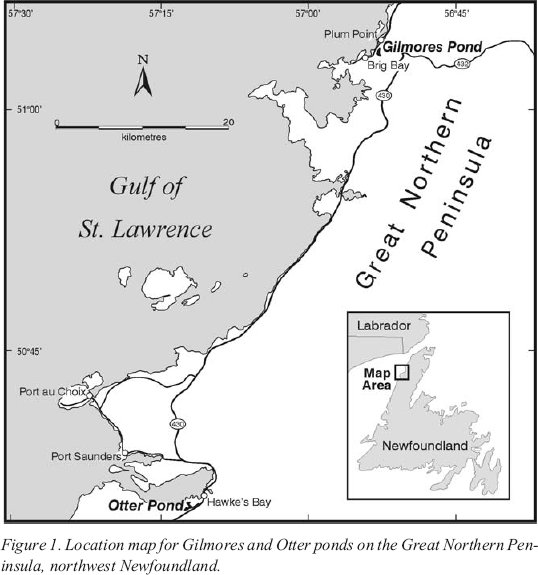
DIATOMS —BIOLOGY AND APPLICATIONS
6 Diatoms are a diverse group of microscopic (5-500 µ#xB5;m), mainly photosynthetic algae (class Bacillariophyceae) that are found almost universally, wherever moisture is present (i.e., lakes, rivers, oceans, hot springs, soil, etc.). The distribution of individual diatom species is controlled by a number of ecological parameters, including acidity, temperature, nutrients, and salinity. Particular to this group of organisms is a cell wall (frustule) composed of silica, which upon death remains well preserved in sediment. Diatom valves have two basic shapes: pennate (linearly and bilaterally symmetrical; Figure 4a) and centric (radially symmetrical; Figure 4b). They may exist in either solitary or colonial forms. The solitary forms are free-living cells that are found floating in water (planktonic forms; Figure 4b), or may be mobile or im-mobile on the basal material. Colonial forms are groups of cells attached to each other by either clasp-like spines or mucilage secretions, or to various plant (epiphytic forms) or mineral substrata (rocks — epilithic; sand grains — episammic; mud — epipelic; Barber and Haworth 1981; Smol 1987). Each diatom frustule is comprised of two separate valves, the morphology of which is unique and identifiable to the species level. Identifications are made with reference to standard taxonomic monographs, such as those of Hustedt (1930) and Patrick and Reimer (1966), as well as numerous smaller, often site-specific studies, i.e., Poulin et al. (1984a). The fact that diatoms can be identified to the species level, and have known ecological and habitat affinities, makes these organisms a strong (though underutilized) tool in archaeological investigations. Examples of diatom applications to archaeology include tracing the provenance of artifacts, assessing the origin, stratigraphy, and depositional environment of archaeological sediments, and reconstructing changes in climate and paleoenvironments (Battarbee 1988; Jug-gins and Cameron 1999). In the present study, however, diatoms are used to reconstruct changes in salinity as a reflection of glacioisostatic uplift and the progressive isolation of basins from open marine conditions to isolated freshwater lakes.
Figure 3. Photograph of raised sea cliff and sea stack behind modern storm beach (covered with driftwood) near Bellburns on the west coast of the Great Northern Peninsula. White arrow points to a person leaning against the sea stack. The raised sea cliff can be observed from many points along the coastal highway (Route 430).Figure 4. (Left) Selected images of diatoms from the two study cores using light microscopy (LM) and scanning electron microscopy (SEM), scale bars of 5 or 10 micrometres (μm) are indicated on each photo: (a) pennate form, Navicula digitoradiata (Greg.) A.Schmidt, x1000 magnification, LM; (b) centric form, Paralia sulcata (Ehrenb.) Cleve, x4300 magnification, SEM; (c) several smaller pennate forms, i) and ii) Amphora pediculus (Kütz) Grun., iii) Fragilaria construens (Ehrenb.) Grun., iv) Fragilaria construens var. venter (Ehrenb.) Hustedt, x1000 magnification, LM; (d) Achnanthes delicatula ssp. hauckiana (Grun.) Lange-Bertalot in Lange-Bertalot and Ruppel 1980, x1250 magnification, LM; (e) Fallacia cryptolyra (Brockman) Stickle and Mann, x10000 magnification, SEM; (f) Cocconeis scutellum var. scutellum Ehrenb., x3300 magnification, SEM.

DIATOM STUDIES OF ISOLATION BASINS
7 One of the strongest controls on diatom distribution is their tolerance of different salinities. Recognizing this trait, diatoms have become a key tool in reconstructing sea-level changes, particularly in formerly glaciated regions, where they mark the change in salinity as a basin became isolated from the sea due to postglacial uplift (Figure 5; Stabell 1987; Pienitz et al. 1991). While the salinity tolerance of some di-atoms is very specific, many diatoms can exist under a range of different salinities, and therefore their distribution may span the breadth of the isolation phase. Thus, while individual species can only give a sense of salinity changes, marked changes in the overall diatom assemblage can be used to collectively estimate the point in a core at which the basin became isolated (or transgressed). This “isolation contact” can then be dated by radiocarbon methods (e.g., Stabell 1987; Pienitz et al. 1991; Denys and de Wolf 1999; Eronen et al. 2001). Isolation of a basin itself can take tens to hundreds of years and is dependent on many factors, including uplift rate and tidal regime. Other considerations include basin and catchment morphometry, marine incursions subsequent to isolation (i.e., storm surges), marine aerosol deposition, and leaching of salts from the surrounding catchment. Thus, recently isolated basins that lie only slightly above sea level can still remain brackish, or retain a lower brackish water layer overlain by freshwater. Such basins are given the name “glo” in Finland, while entirely freshwater basins are called “flada” (Munsterhjelm 1987). Diatom assemblages from glo basins can be expected to exhibit both freshwater and brackish floras.
Figure 5. An illustration of how coring formerly submerged lakes might resolve changes in relative sea level, including possible short-lived changes. The curve on the right illustrates the case of an emergent coastline that undergoes a late, low amplitude sea-level oscillation wherein the low lying coastal areas are isolated, then are transgressed by a relative rise in sea level (submerged) and then are once again isolated through relative sea-level regression. The figure at left illustrates the expected sediment sequence one would find when coring higher elevation lakes (Lake A) wherein a single lake basin isolation phase occurs. In contrast, low lying basins within the amplitude of the late phase relative sea-level oscillation (Lake B) would be expected to record a sequence of marine-brackish-fresh-brackish-marine-brackish-freshwater organisms. The diatom sampling strategy employed in this study (visual inspection of samples every 5 cm core depth, and detailed inspections every 1 cm in the upper 10-cm core depth) would be able to detect both isolation/transgression scenarios if present in the study cores.
SALINITY CLASSIFICATION
8 In a very generalized fashion, the halobian, or salinity, classification for diatoms can be broken down into four categories: (i) Halophobous [strictly freshwater], 0l (parts per thousand) salt, (ii) Oligohalobous (indifferent (ind.) or halophilic) [fresh-weakly brackish], .0.2l salt, (iii) Mesohalobous [brackish], ~30 to 0.2l salt, and (iv) Polyhalobous [marine], /30l salt. Of these three, the mesohalobous, or brackish, phase is the most difficult to define in both a literal and numerical sense, as any divisions are sure to be artificial rather than defined according to biological thresholds. While several halobian classification schemes have been developed, those that combine salinity ranges with habitat, or life forms, seem to provide the greatest insight into past environmental changes, and have thus been utilized here.
STUDY LAKE SELECTION AND CORING
9 In order to preserve a record of recent marine submergence by a sea-level rise of approximately 3-4 metres, a lake basin threshold must lie between the present shoreline and at most 3 metres above sea level. Candidate basins in the Port au Choix region that tentatively met this requirement were identified on aerial photographs and 1:50,000 topographic maps (10 m contour interval), based on their close proximity to the present coastline or their situation at elevations below 10 metres above sea level, respectively. Lake basins located near coastal towns were checked on 1:2500 or 1:5000 community maps (contour interval 2 m), on which surveyed lake levels were provided. Summer field reconnaissance eliminated lakes dammed by beach berms and extremely shallow or deep lakes, where sediment thicknesses were limited or sediment reworking was likely. Lake outlets were also examined and surveyed.
10 Two lakes met the criteria for sampling. Otter Pond is located within the town limits of Hawke’s Bay at 1.5± metres above sea level and within 450 metres of the present coastline (Figure 1). The lake is approximately 50 hectares in area and mostly 1 metre or less in depth. The coring location (50.592EN; 57.195EW) was 90 metres out from the midpoint of the southern shoreline in 1.3-metre water depth. Historically, the lake was used to store logs for the spring drive through the outlet channel to Hawkes Bay. As a result, the outlet channel has been straightened but there is no apparent evidence for deepening of the channel, which is floored by large boulders.
11 Gilmores Pond is the local name for a 25-hectare lake located between the communities of Brig Bay and Plum Point (51.062EN; 56.891EW; Figure 1). The lake outlet is at an elevation of approximately 0.3 metres above sea level and is separated from Brig Bay by a broad shallow inlet (500 m long) that is inundated by the sea during high water levels. The coastal road crosses the outlet and bedrock is encountered at the surface nearby, suggesting that the basin is a bedrock hollow. The lake is more than 10 metres deep, with the deepest area near its southeast corner. The community of Brig Bay had considered using the lake as a domestic water source but encountered brackish water at the bottom (indicating this is a glo basin).
12 Coring was carried out in the winter, enabling the lake ice to be used as a coring platform. A hole was drilled through the ice cover and then sediment cores were extracted from the deepest basin of each pond, using a simple percussion corer (Reasoner 1993). Retrieved core lengths were 1.8 metres in Gilmores Pond and 2.4 metres in Otter Pond. Core tubes were capped and sealed in the field, and then returned to the laboratory where they were split lengthwise and subsampled for diatoms and other sedimentological and biological characteristics.
SAMPLE PREPARATION AND ANALYSIS
13 Following a visual description of the sediment cores, samples from halved sections were taken every 10 centimetres for bulk density, organic and carbonate content, and every 5 centimetres for diatom identification. Sediment colour was recorded using a Munsell soil colour chart. Standard analytical procedures using loss-on-ignition (LOI) methods were followed for bulk density, and organic and carbonate content (Berglund 1986). Samples (1 cc) for diatom analysis were first subjected to a digestion in 10% hydrochloric acid (HCl) to remove carbonates, and then 30% hydrogen peroxide (H2O2) to remove organic matter. Cleaned samples were then dispersed onto coverslips and mounted onto glass slides using Meltmount8(refractive index = 1.704) according to standard diatom procedures (Battarbee 1986). A total of 500 diatom valves were counted on each slide under oil immersion at x1250 magnification.
14 Species identification and salinity classification was made with reference to Bérard-Therriault et al. (1986, 1987), Campeau et al. (1999), Cardinal et al. (1984, 1986), Foged (1978), Krammer and Lange-Bertalot (1986, 1988, 1991a,b), Mölder and Tynni (1969, 1972), Peragallo and Peragallo (1897-1908), Poulin et al. (1984a,b,c), van Dam et al. (1994), and Vos and de Wolf (1988, 1993). Fragilaria nomenclature sensu Hustedt (1930) and Patrick and Reimer (1966) is used in this study.
SEDIMENT CORE STRATIGRAPHY
Gilmores Pond
15 The Gilmores Pond core consists of light grey silty-clay with pebbles, organic macrofossils and bands of coarse silt and fine sand (174-171 cm), silty clay (171-130 cm) clayey-silt (130-44 cm) and silty-gyttja (44-0 cm; Figure 6). The percussion corer was unable to penetrate the lowermost gritty layer. Light grey sand lenses occur intermittently through the core, starting at the lower contact of the clayey-silt unit (130 cm), while fine gravel and single clasts were either dispersed through the matrix or concentrated in discrete intervals (e.g., 139-144 cm). The possible remains of a shell was present at 110 centimetres, foraminifera were abundant between 146 and 148 centimetres, and plant macrofossils occurred throughout, although discrete concentrations were also observed (40 cm, 53-58 cm, 74-94 cm, 99-105 cm, 139-144 cm). There was a distinct fissility to the sediment below 150 centimetres and the upper 5 centimetres of sediment appeared mixed.
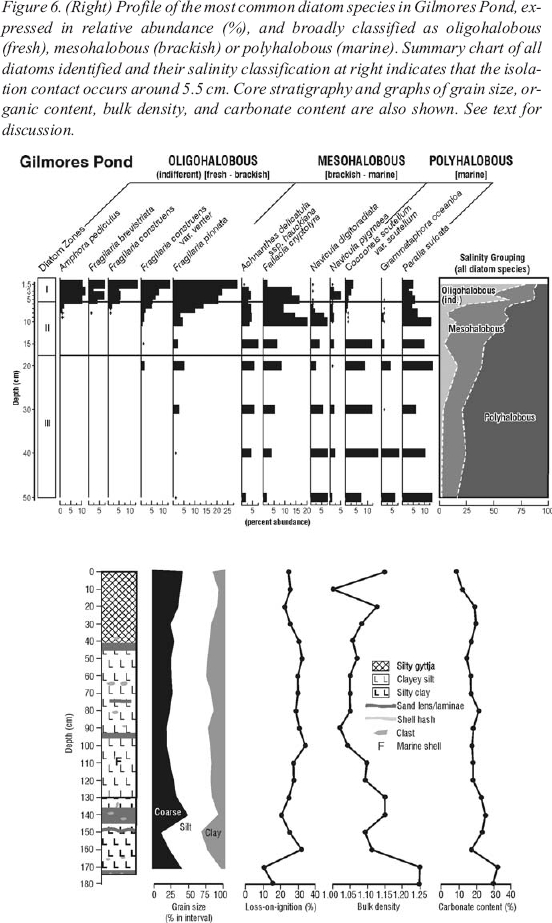
Figure 6. (Right) Profile of the most common diatom species in Gilmores Pond, expressed in relative abundance (%), and broadly classified as oligohalobous (fresh), mesohalobous (brackish) or polyhalobous (marine). Summary chart of all diatoms identified and their salinity classification at right indicates that the isolation contact occurs around 5.5 cm. Core stratigraphy and graphs of grain size, organic content, bulk density, and carbonate content are also shown. See text for discussion.
Display large image of Figure 6
16 The carbonate content of the core varies between 9 and 20%, rising to 30% in the lowest 20 centimetres. Bulk density readings decrease from 1.13 at the core top to 1.03 at 90 centimetres, then increase to 1.15 at 140 centimetres, finally reaching1.25 at the core bottom. The organic content is between 20 and 30% for much of the core but decreases by half in the lower 10 centimetres or so. Textural analysis shows gradual fining downcore from approximately 40% sand and fine gravel at the top to 20% at approximately 100 centimetres. Below 100 centimetres, the texture remains reasonably consistent except for one coarse (up to 50% sand or coarser) and one fine layer (95% silt and clay) between 135 and 150 centimetres.
17 The relative consistency of the various sedimentological parameters measured in the Gilmores Pond core above 140 centimetres depth likely reflects the pond’s proximity to the open sea, and a relatively constant flux of sediments from more or less the same catchment area.
Otter Pond
18 The Otter Pond core consists of a black silty gyttja in its upper 28 centimetres overlying very dark grey or greyish brown clayey silt to 130 centimetres (Figure 7). The contact is marked by a thin layer of shell hash and a 1.5-centimetre-thick, grey, well-sorted fine to medium sand layer. Below 130 centimetres, the sediment texture is grittier and there is a slight colour change to very dark greyish brown. Whole shells, shell fragments and shell hash layers occur infrequently below this level. Single clasts are rare between 28 and 62 centimetres, but more common from 62 to 155 centimetres. Lenses of fine to medium dark grey sand, up to 1.5 centimetres thick, occur between 129 centimetres and the base of the core, with numerous layers below 171 centimetres. The lenses appear multi-banded with finer interbeds visible in some. Organics occur throughout the core but are more concentrated in the upper 18-20 centimetres. Fissility appears in the core below 85 centimetres, possibly as a reflection of the sediment texture.
Figure 7. (Right) Profile of the most common diatom species in Otter Pond, expressed in relative abundance (%), and broadly classified as oligohalobous (fresh), mesohalobous (brackish) or polyhalobous (marine). Summary chart of all diatoms identified and their salinity classification at right indicates that the isolation contact occurs around 1 cm. Core stratigraphy and graphs of grain size, organic content, bulk density, and carbonate content are also shown. See text for discussion.

Display large image of Figure 7
19
19 The Otter Pond core has a fairly uniform carbonate content downcore to 100 centimetres, varying between 5.5 and 6%. By 125 centimetres depth it has decreased to 2-3.5% but shows greater variability in the lower half of the core. The bulk density in the upper 30 centimetres decreases from 1.2 to 1 and then remains fairly stable to 120 centimetres depth. Below 120 centimetres it fluctuates between1.25 and 1.5. The organic content of the core increases from 5% at the top to 15% at 90 centimetres and then progressively decreases to 2-5% at the core bottom. The top of the core appears relatively coarse with 50-65% sediment coarser than silt and 15% or less as clay. By 80 centimetres depth the coarse fraction has decreased to 25% and the clay fraction has increased to 35%. Between 100 and 170 centimetres depth there is 50-70% coarse and 10% or less clay. Below 170 centimetres, sediment texture is more variable with fluctuations in the coarse fraction between 35 and 70% and corresponding changes in clay. The variability in texture, bulk density, and carbonate and organic contents through the lowest 70 centimetres of the core likely reflects sampling of discrete sand lenses, which appear more abundant in the lower third of the core. The same pattern is also seen at 20 centimetres depth in the core where sampling of a sand lens resulted in a peak in the coarse sand fraction, a decrease in carbonate and organic contents, and an increase in bulk density.
20 The measured sedimentological properties of the Otter Pond core appear to record a prominent shift around 120 centimetres depth where bulk density decreases and organic content (measured by LOI) and carbonate content correspondingly increase. This likely reflects progressive emergence of the coastline around the pond, isolation from the open ocean (the entrenchment of Hawke’s Bay), and a shift in dominance to more local terrestrial sediment sources.
DIATOM STRATIGRAPHIES
21 Visual scans at x1250 magnification of prepared diatom samples (visually assessed every 5 cm and counted every 10-cm interval) revealed that the sediments of the two ponds were dominated by marine and brackish flora for almost their entire length. Freshwater (oligohalobous (ind.)) taxa were only prominent in the uppermost (<10 cm) sediment, wherein the cores were subsampled and diatoms counted at 1-centimetre intervals. Halophobous taxa were collectively very rare (<1%), which likely reflects the recent emergence of these basins and their proximity to the sea. Diatom abundance was high in all samples, and while counts from each slide commonly contained more than 40 different species, most individual species were too sparse to be meaningfully represented. Percentage abundances of the dominant species in Gilmores and Otter ponds are presented in Figures 6 and 7.
Gilmores Pond
22 The uppermost 7 centimetres of sediment of the Gilmores Pond core is dominated by oligohalobous (ind.) Amphora pediculus (Kütz) Grun. and Fragilaria spp., notably Fragilaria pinnata Ehrenb. and Fragilaria construens var. venter (Ehrenb.) Grun. (Figures 4c and 6). A similar dominance of Fragilaria taxa in recently emerged basins was noted by Stabell (1985) in studies of Scandinavian lakes and ponds. Each of the four Fragilaria species are tychoplanktonic taxa, meaning that they live both within the benthic regions and suspended in the water column (by currents and wind/wave action). Below the oligohalobous (ind.) dominated core section, a number of epipelic (unattached, motile in and on sediments) and epipsammic (attached to sand grains) diatoms are prominent. Achnanthes delicatula ssp. hauckiana (Grun.) Lange-Bertalot (Figure 4d) is an epipsammic form that was noted by Witkowski (1991) and Campeau et al. (1999) as dominating sandy shallow coastal environments. Fallacia cryptolyra (Brockman) Stickle and Mann (Figure 4e) is an epipelic form reported from Atlantic coastal sediments (Pienitz et al. 1991). Other mesohalobous taxa seen in this core (Navicula digitoradiata (Greg.) A. Schmidt (Figure 4a) and Navicula pygmaea Kütz) are commonly reported in coastal and basin isolation studies as shallow water brackish forms (e.g., Riznyk 1973; Long et al. 1999). The increase in polyhalobous taxa below10 centimetres in the core indicates a much stronger marine signal. Prominent species include the marine-brackish epiphytes (living attached to plants) Cocconeis scutellum var. scutellum Ehrenb. (Figure 4f) and Grammataphora oceanica Ehrenb., and the marine-planktonic (free-floating in the water column) Paralia sulcata (Ehrenb.) Cleve (Figure 4b). Studies have suggested that owing to their ecology, dispersion and the robustness of the valves, species such as P. sulcata may become falsely enriched in the thanatocoenoses (fossil assemblage — as opposed to actual living assemblages; Vos and de Wolf 1993; Denys 1994). This may account for their relatively high percentages in the uppermost, oligohalobous (ind.) dominated sediment.
23 Based on the complete diatom assemblage from this core, the isolation contact is placed at approximately 5.5 centimetres depth, coincident with a marked increase in percentage abundance and diversity of oligohalobous (ind.) taxa (Figure6). Continued presence of marine and brackish water taxa in the uppermost sediments likely signals the reworking of raised marine sediments, or possibly, continued incursions of seawater into this basin. An age of 600 cal BP (Table 1) was determined on unidentified plant material from 2-4 centimetres depth in the core and provides a minimum date on the isolation contact.
Otter Pond
24 The diatom stratigraphy of Otter Pond is dominated (25-65%) by the marine-brackish epiphyte Cocconeis scutellum var. scutellum Ehrenb. (Figures 4f and7). This species is commonly reported in shallow, marine benthic samples from the Maritimes (Bacon and Taylor 1976; Poulin et al. 1984a) and in similar isolation basin studies on Greenland (Long et al. 1999). Other prominent epiphytic species include the brackish-marine Cocconeis costata Greg., Cocconeis scutellum var. parva (Grun. in Van Heurck) Cleve and Tabularia fasciculata (Agardh) Williams and Round 1986. Most of the other brackish and marine species are epipelic or epipsammic. Paralia sulcata (Ehrenb.) Cleve (Figure 4b) is a marine planktonic form, which in the Maritimes is commonly associated with nutrient-rich waters (Bacon and Taylor 1976; Bérard-Therriault et al. 1987; Lapointe 2000). The oligohalobous (ind.) Fragilaria taxa that dominate the upper sediment of Gilmores Pond only occur at low percentage abundances (<10%) in the upper 3 centimetres of Otter Pond. This suggests that Otter Pond may only recently have emerged from the sea, that the uppermost sediment was lost during coring, or that the uppermost sediment has been disturbed by human activities (the pond was previously dammed to facilitate the spring flushing of timber to the sea).
25 Tentatively, the isolation contact is placed around 1 centimetre depth, although the initial uplift of the outlet may be signaled by the first rise in oligohalobous (ind.) taxa around 3 centimetres. An age of 130 cal BP (Table 1) on Picea needles from 5.5 centimetres depth in the core provides a maximum on initial isolation of Otter Pond from the sea.
Table 1. Details of radiocarbon-dated samples from Gilmores and Otter ponds

DISCUSSION
26 The diatom records presented from Gilmores and Otter ponds (Figures 6 and 7), and visual scans of samples at 5-centimetre intervals from throughout the total core lengths, indicate that these coastal basins only recently became isolated from the sea. Despite the absence of basal radiocarbon dates that would conclusively demonstrate the lengths of record in each core, we are confident that in consideration of low sedimentation rates recorded in similar basins and environmental settings (e.g., Pienitz et al. 1991; Anderson and Macpherson 1994), both cores extend into the early Holocene, and hence would incorporate a late Holocene sea-level oscillation had one occurred. Further evidence in support of this assertion is the observation in both cores, that with decreasing core depth, there is a progressive increase in epiphytic diatom taxa and an accordant decrease in epipelic and epipsammic species. This likely indicates an increase in the amount of plant material, and would be expected as the basin was raised further into the optimal photic zone. Similarly, the sedimentology and marine macrofossil records (Figures 6 and 7) support the assertion of progressive uplift of marine basins and emergence only in the uppermost sediment. Thus, the pattern of progressive shallowing and absence of oligohalobous (ind.) diatom taxa throughout all but the uppermost sediment is fully in agreement with the notion that these basins were being uplifted under a type-A sea-level history, and therefore disputes Grant’s (1994) notion of a late Holocene sea-level oscillation, or modified type-B curve.
27 If the horizontal sea cliff along the west coast of the Great Northern Peninsula is therefore not of late Holocene age, when was it formed? One possible answer is that it dates to the last interglacial period about 120,000 years ago. Assuming there have been no net changes in crustal position or oceanographic conditions over the last glacial period, then the last interglacial shore should be coincident or parallel to the present shore. For example, a relic shore platform has been observed fairly consistently at about 4-6 metres above its modern counterpart on the Burin Peninsula, the Avalon Peninsula, and parts of southwest Newfoundland (Grant 1989). A similar feature and related deposits elsewhere in Atlantic Canada have been attributed to the last interglacial (Grant 1989), and therefore it is not unreasonable to consider the raised sea cliff on the Great Northern Peninsula a northeastern extension of this interglacial shore.
28 On the basis of the confirmed sea-level history and reconstructed palaeoshoreline configuration for Port au Choix, we were able to establish that the MAI cemetery was located on an ancient island, and to identify a prominent raised marine terrace between 6 and 8 metres above sea level, on what would have been a sheltered “mainland” shoreline, as the strongest candidate for a MAI living site. We selected the most likely location for MAI habitation on this ancient shoreline by identifying critical site selection factors such as proximity to fresh water, accessibility by boat, a well-drained substrate, and a flat broad terrace. Employing this site prediction strategy, intensive and sustained trenching through 1.4-metre-thick peat in a wooded area was conducted, and eventually yielded a MAI non-mortuary site (the Gould Site, EeBi-42, Renouf and Bell 2002). Without this study’s integrated assessment of sea-level history, coastal geomorphology, and archaeology, it is unlikely that we would have otherwise persisted with such a time-consuming testing programme, and achieved the success that we did.
29 Having demonstrated the utility of the isolation basin approach for testing sea-level histories in the Port au Choix area, we can project its potential use elsewhere in Newfoundland and Labrador in the following circumstances: Where raised marine sediments and organic remains suitable for radiocarbon dating are either absent or poorly preserved. Much of coastal Labrador is proposed as a continuously emergent landscape (type-A sea-level history; Clark and Fitzhugh 1992), but the bedrock-dominated coastline rarely preserves raised marine deposits, except in sheltered inlets and estuaries (e.g., Porcupine Strand, Smith et al. 2003), and carbonate fossils (e.g., shells) are commonly dissolved in the acidic groundwater.
30 RSL curves therefore are poorly constrained and rely on modeled uplift parameters (e.g., exponential decay constant “k,” Clark and Fitzhugh 1992). Numerous rock basins below marine limit, however, could potentially preserve a marine-lacustrine isolation sequence that could be radiocarbon dated using plant macrofossils (see, for example, Jordan 1975). Selective application of the isolation basin approach would thus generate sea-level data in support of archaeological investigations, and further constrain modeled sea-level history.
31 Where much of the Holocene sea-level record is submerged in the shallow offshore. For most of the Island of Newfoundland, sea level has been rising from an early to mid Holocene lowstand. The depth of the lowstand generally decreases towards the interior and the Great Northern Peninsula, with maximum values (-30 m) off the south and southwest coasts. The lowstand sea level has rarely been dated, but is likely to have occurred earlier in the offshore and outer coast than in nearshore areas. Bell and Renouf (2003) argued that this variable and complex sea-level history caused the uneven coastal distribution of late MAI sites (6290-3340 cal BP), and the apparent absence of early MAI sites (8860-6290 cal BP), despite their presence in nearby southern Labrador. They proposed that in areas where the lowstand is greater than -20 metres water depth, late MAI sites older than 3200 cal BP are today submerged, whereas in all areas outside the Great Northern Peninsula (which has experienced continuous emergence), early MAI sites would similarly be under water.
32 Testing this hypothesis initially requires detailed sea-level reconstruction from submerged records of former coastal environments. Unfortunately, the marine transgression across these environments can rework and destroy the landform and sedimentary evidence. One exception is the sedimentary record preserved in transgression basins, which acted as freshwater lakes when sea level was lower, but subsequently were transgressed by salt water as sea level rose. Such basins have been identified in the nearshore off northeast Newfoundland and are the focus of current coring initiatives.
Acknowledgements
Initial and substantial support for this project was provided by a New Initiatives Research Grant, Office of the Vice President (Research), Memorial University of Newfoundland, to Trevor Bell and M.A.P. Renouf. The Natural Sciences and Engineering Research Council of Canada provided subsequent funding to Trevor Bell. Rod Smith was originally supported in this research by Athabasca University, and subsequently by the Geological Survey of Canada (Natural Resources Canada ESS contribution #2004302). We would like to thank our coring crew: Tim Rast, Naomi Short, Ralph House, and Todd Chislett. Kevin Sheppard performed sediment analysis. David Mercer, Memorial University of Newfoundland Cartography Laboratory, drafted diagrams. We acknowledge the constructive comments of two reviewers.References
Anderson, T.W. and J.B. Macpherson (1994), “Wisconsinan Late-glacial environmental change in Newfoundland: a regional review.” Journal of Quaternary Science 9: 171-178.
Bacon, G.B. and A.R.A. Taylor (1976), “Succession and stratification in benthic diatom communities colonizing plastic collectors in a Prince Edward Island estuary.” Botanica Marina 19: 231-240.
Barber, H.G. and E.Y. Haworth (1981), A Guide to the Morphology of The Diatom Frustule: with a key to the British freshwater genera. Freshwater Biological Association Scientific Publication no. 44.
Barnhardt, W.A., W.R. Gehrels, D.F. Belknap, and J.T. Kelley (1995), “Late Quaternary relative sea-level change in the western Gulf of Maine: evidence for a migrating glacial forebulge.” Geology 23: 317-320.
Battarbee, R.W. (1986), “Diatom analysis.” In Handbook of Holocene Palaeoecology and Palaeohydrology, edited by B.E. Berglund, pp. 527-570. Toronto: John Wiley & Sons.
Battarbee, R.W. (1988), “The use of diatom analysis in archeology: A review.” Journal of Archeological Science 15: 621-644.
Bell, T. and M.A.P. Renouf (1998). “Integrating Archaeology and Sea Level History at Port au Choix, NW Newfoundland.” In Assessing the State of the Environment of Gros Morne National Park, edited by D.W. Anions and A.R. Berger, pp. 109-113. Parks Canada — Ecosystem Science Review Report 011.
Bell, T. and M.A.P. Renouf (2003), “Prehistoric Cultures, Reconstructed Coasts: Maritime Archaic Indian Site Distribution in Newfoundland.” World Archaeology 35: 350-370.
Bell, T., D.G.E. Liverman, M.J. Batterson, and K. Sheppard (2001), “Late Wisconsinan Stratigraphy and Chronology of Southern St. George’s Bay, Southwest Newfoundland: a Re-appraisal.” Canadian Journal of Earth Sciences 38: 851-869.
Bell, T., I.R. Smith, and M.A.P. Renouf (this volume), “Postglacial Sea-Level History and Coastline Change at Port au Choix, Great Northern Peninsula, Newfoundland.” Bérard-Therriault, L., A. Cardinal, and M. Poulin (1986), “Les Diatomées (Bacillariophyceae) benthiques de substrats durs des eaux marines et saumâtres du Québec. 6. Naviculales: Cymbellaceae et Gomphonemaceae.” Naturaliste Canadien 113: 405-429.
Bérard-Therriault, L., A. Cardinal, and M. Poulin (1987), “Les Diatomées (Bacillariophyceae) benthiques de substrats durs des eaux marines et saumâtres du Québec. 8. Centrales.” Naturaliste Canadien 114: 81-103.
Berglund, B.E. (1986), Handbook of Holocene Palaeoecology and Palaeohydrology. Toronto: John Wiley & Sons, 869 p.
Campeau, S., R. Pienitz, and A. Héquette (1999), “Diatoms from the Beaufort Sea coast, southern Arctic Ocean (Canada).” Bibliotheca Diatomologica, Band 42. Berlin: J. Cramer, 244 p.
Cardinal, A., M. Poulin, and L. Bérard-Therriault (1984), “Les Diatomées benthiques de substrats durs des eaux marines et saumâtres du Québec. 4. Naviculales, Naviculaceae (à l’exclusion des genres Navicula, Donkinia, Gyrosigma et Pleurosigma).” Naturaliste Canadien 111: 369-394.
Cardinal, A., M. Poulin, and L. Bérard-Therriault (1986), “Les Diatomées benthiques de substrats durs des eaux marines et saumâtres du Québec. 5. Naviculales, Naviculaceae; les genres Donkinia, Gyrosigma et Pleurosigma.” Naturaliste Canadien 113: 167-190.
Clark, P.U. and W.W. Fitzhugh (1992), “Postglacial Relative Sea Level History of the Labrador Coast and Interpretation of the Archaeological Record.” In Paleoshorelines and Prehistory: an Investigation of Method, edited by L.L. Johnson, pp. 189-213. London: CRC Press.
Corner, G.D., V.Y. Yevzerov, V.V. Kolka, and J.J. Moller (1999), “Isolation basin stratigraphy and Holocene relative sea-level change at the Norwegian-Russian border north of Nikel, northwest Russia.” Boreas 28: 146-166.
Denys, L. (1994), “Diatom assemblages along a former intertidal gradient: A paleoecological study of a Subboreal clay layer (western coastal plain, Belgium).” Netherlands Journal of Aquatic Ecology 28: 85-96.
Denys, L. and H. de Wolf (1999), “Diatoms as indicators of coastal paleo-environments and relative sea-level change.” In The Diatoms: Applications for the Environmental and Earth Sciences, edited by E.F. Stoermer and J.P. Smol, pp. 277-297. Cambridge: Cambridge University Press.
Eronen, M., G. Glückert, L. Hatakka, O. van de Plassche, J. van der Plicht, and P. Rantala (2001), “Rates of Holocene isostatic uplift and relative sea-level lowering of the Baltic in SW Finland based on studies of isolation contacts.” Boreas 30: 17-30.
Foged, N. (1978), Diatom Analyses. The Archaeology of Svendborg, Denmark. No. 1.Odense University Press.
Grant, D.R. (1989), “Quaternary geology of the Appalachian region of Canada.” In Quaternary Geology of Canada and Greenland, edited by R.J. Fulton, pp. 393-440. Ottawa: Geological Survey of Canada.
Grant, D.R. (1994), “Quaternary Geology of Port Saunders Map Area, Newfoundland.”Geological Survey of Canada, Paper 91-20.
Hustedt, F. (1930), “Bacillariophyta (Diatomaceae).” In Die Süsswaser-floraMitteleuropas. Heft 10, edited by A. Pascher. Jena: G. Fischer.
Hutchinson, I., T. James, J. Clague, J.V. Barrie, and K. Conway (2004), “Reconstruction of late Quaternary sea-level change in southwestern British Columbia from sediments in isolation basins.” Boreas 33: 183-194.
Jordan, R. (1975), “Pollen diagrams from Hamilton Inlet, central Labrador and their environmental implications for the northern Maritime Archaic.” Arctic Anthropology 12: 92-116.
Josenhans, H., D. Fedje, R. Pienitz, and J. Southon (1997), “Early Humans and Rapidly Changing Holocene Sea Levels in the Queen Charlotte Islands — Hecate Strait, British Columbia, Canada.” Science 277: 71-74.
Juggins, S. and N. Cameron (1999), “Diatoms and archaeology.” In The Diatoms: Applications for the Environmental and Earth Sciences, edited by E.F. Stoermer and J.P. Smol, pp. 389-401. Cambridge: Cambridge University Press.
Krammer, K. and H. Lange-Bertalot (1986), “Bacillariophyceae. 1. Teil: Naviculaceae.” In Süßwasserflora von Mitteleuropa, Band 2/1, edited by H. Ettl, G. Gärtner, J. Gerloff,H. Heynig, and D. Mollenhauer. New York: Gustav Fisher Verlag.
Krammer, K. and H. Lange-Bertalot (1988), “Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiacea, Surirellaceae.” In Süßwasserflora von Mitteleuropa, Band 2/2, edited by H. Ettl, G. Gärtner, J. Gerloff, H. Heynig, and D. Mollenhauer. New York: Gustav Fisher Verlag.
Krammer, K. and H. Lange-Bertalot (1991a), “Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae.” In Süßwasserflora von Mitteleuropa, Band 2/3, edited by H. Ettl, G. Gärtner, J. Gerloff, H. Heynig, and D. Mollenhauer. Stuttgart: Gustav Fisher Verlag.
Krammer, K. and H. Lange-Bertalot (1991b), “Bacillariophyceae. 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema Gesamtliteraturverzeichnis.” In Süßwasserflora von Mitteleuropa, Band 2/4, edited by H. Ettl, G. Gärtner, J. Gerloff, H. Heynig, and D. Mollenhauer. Stuttgart: Gustav Fisher Verlag.
Lapointe, M. (2000), “Modern diatom assemblages in surface sediments from the Maritime Estuary and the Gulf of St. Lawrence, Québec (Canada).” Marine Micropaleontology 40: 43-65.
Long, A.J., D.H. Roberts, and M.R. Wright (1999), “Isolation basin stratigraphy and Holocene relative sea-level change on Arveprinsen Ejland, Disko Bugt, West Greenland.” Journal of Quaternary Science 14: 323-345.
Mölder, K. and R. Tynni (1969), “Über Finnlands rezente und subfossile diatomeen III.” Bulletin Geological Society of Finland 41: 235-251.
Mölder, K. and R. Tynni (1972), “Über Finnlands rezente und subfossile diatomeen VI.” Bulletin Geological Society of Finland 44: 141-149.
Mörner, N.-A. (1976), “Eustasy and geoid changes.” The Journal of Geology 84: 123-151. Mörner, N.-A. (1992), “Sea-level change and Earth’s rate of rotation.” Journal of Coastal Research 8: 966-971.
Munsterhjelm, R. (1987), “Flads and gloes in the archipelago. Fennoscandian Land Uplift.” Proceedings of a symposium at Tvärminne, 10-11 April 1986. Geological Survey of Finland Special Paper 2: 55-61.
Patrick, R. and C.W. Reimer (1966), “The diatoms of the United States exclusive of Alaska and Hawaii. Vol. 1.” Philadelphia: Academy of Natural Sciences, Monograph 13. Peragallo, H. and M. Peragallo (1897-1908), “Diatomées marines de France et des districts maritimes voisins.” Micrographe-Éditeur, à Grez-sur-Loing (S.-et-M.). 491 pp., Atlas, 137 pp.
Pienitz, R., G. Lortie, and M. Allard (1991), “Isolation of lacustrine basins and marine regression in the Kuujjuaq area, northern Québec, as inferred from diatom analysis.” Géographie physique et Quaternaire 45: 155-174.
Poulin, M., L. Bérard-Therriault, and A. Cardinal (1984a), “Les Diatomées benthiques de substrats durs des eaux marines et saumâtres du Québec 1. Cocconeioideae (Achnanthales, Achnanthaceae).” Naturaliste Canadien 111: 45-61.
Poulin, M., L. Bérard-Therriault, and A. Cardinal (1984b), “Les Diatomées (Bacillariophyceae) benthiques de substrats durs des eaux marines et saumâtres du Québec 2. Tabellarioideae et diatomoideae (Fragilariales, Fragilariaceae).” Naturaliste Canadien 111: 275-295.
Poulin, M., L. Bérard-Therriault, and A. Cardinal (1984c), “Les Diatomées benthiques de substrats durs des eaux marines et saumâtres du Québec 3. Fragilarioideae (Fragilariales, Fragilariaceae).” Naturaliste Canadien 111: 349-367.
Quinlan, G. and C. Beaumont (1981), “A Comparison of Observed and Theoretical Postglacial Relative Sea Level in Atlantic Canada.” Canadian Journal of Earth Sciences 19: 1146-1163.
Reasoner, M.A. (1993), “Equipment and procedure improvements for a lightweight, inexpensive, percussion core sampling system.” Journal of Paleolimnology 8: 273-281.
Renouf, M.A.P. and T. Bell (2002), “Integrating Sea Level History and Geomorphology in Targeted Archaeological Site Survey: The Gould Site (EeBi-42), Port au Choix, Newfoundland.” Northeastern Anthropology 59: 47-64.
Renouf, M.A.P. and T. Bell (in press), “Maritime Archaic Site Locations on the Island of Newfoundland.” In The Archaic of the Far Northeast, edited by D. Sanger and M.A.P. Renouf. Orono, ME: University of Maine Press.
Riznyk, R.Z. (1973), “Interstitial diatoms from two tidal flats in Yaquina Estuary, Oregon,USA.” Botanica Marina 16: 113-138.
Sanger, D. and D.F. Belknap (1987), “Human Responses to Changing Marine Environments in the Gulf of Maine.” In Man and the Mid-Holocene Climatic Optimum: Proceedings of the 17th Annual Chacmool Conference, edited by N. McKinnon and G. Stuart, pp. 245-261. Department of Archaeology, University of Calgary, Calgary.
Sanger, D. and D.L. Kellogg (1989), “Prehistoric Archaeology and Evidence of Coastal Subsidence on the Coast of Maine.” In Neotectonics of Maine, edited by W.A. Anderson and H.W. Borns Jr., pp. 107-126. Maine Geological Survey, Augusta.
Scott, D.B. and E.S. Collins (1996), “Late Mid-Holocene Sea-level Oscillation: A PossibleCause.” Quaternary Science Reviews 15: 851-856.
Smith, J., T. Bell, and L. Rankin (2003), “Quaternary Geology and Landscape Change, Porcupine Strand, Labrador.” Newfoundland Department of Mines and Energy, Geological Survey Report 03-1: 293-305.
Smol, J.P. (1987), “Methods in Quaternary Ecology #1. Freshwater Algae.” GeoscienceCanada 14: 208-217.
Stabell, B. (1985), “The development and succession of taxa within the diatom genus Fragilaria Lyngbye as a response to basin isolation from the sea.” Boreas 14: 273-286.
Stabell, B. (1987), “Changes in diatom floras in late Quaternary Western and Southeastern Norwegian marine and freshwater sediments: Response to basin isolation from the sea.” Nova Hedwigia 44: 305-326.
Stuiver, M. and P.J. Reimer (1993), “Extended 14C database and revised CALIB radiocarbon calibration program.” Radiocarbon 35: 215-230.
Stuiver, M., P.J. Reimer, E. Bard, W.E. Beck, G.S. Burr, K.A. Hughen, B. Kromer, F.G. McCormac, J. van der Plicht, and M. Spurk (1998), “INTCAL98 radiocarbon age calibration 0-24,000 BP.” Radiocarbon 40: 1041-1083.
Tuck, J.A. (1976), Ancient Peoples of Port au Choix Newfoundland. Institute of Social and Economic Research, Social and Economic Studies 17, Memorial University of Newfoundland, St. John’s.van Dam, H., A. Mertens, and J. Sinkeldam (1994), “A coded checklist and ecological indicator values of freshwater diatoms from The Netherlands.” Netherlands Journal of Aquatic Ecology 28: 117-133.
Vos, P.C. and H. de Wolf (1988), “Methodological aspects of paleo-ecological diatom research in coastal areas of the Netherlands.” Geologie en Mijnbouw 67: 31-40.
Vos, P.C. and H. de Wolf (1993), “Diatoms as a tool for reconstructing sedimentary environments in coastal wetlands; methodological aspects.” Hydrobiologia 269/270: 285-296.
Witkowski, A. (1991), “Diatoms of the Puck Bay coastal shallows (Poland, Southern Baltic).” Nordic Journal of Botany 11: 689-701.
Notes