Series
Great Canadian Lagerstätten 3.
Late Ordovician Konservat-Lagerstätten in Manitoba
SUMMARY
Konservat-Lagerstätten, deposits in which soft-bodied or lightly sclerotized fossils are preserved, are very rare in Ordovician strata. Three significant sites are known from Upper Ordovician rocks in Manitoba: at Cat Head – McBeth Point, William Lake, and Airport Cove. These sites are in two distinct sedimentary basins: the former two are in the Williston Basin, while the latter is in the Hudson Bay Basin. All three sites contain marine fossils, but each has a different assemblage that contributes a distinct piece of the diversity picture. Important groups represented at one or more of the sites include seaweeds (algae), sponges, cnidarian medusae (jellyfish), conulariids, trilobites, eurypterids, xiphosurids (horseshoe crabs), and pycnogonids (‘sea spiders’). The different biotas reflect depositional conditions at each site. Many of the fossils are unknown elsewhere in the Ordovician at the family level or higher. The province of Manitoba therefore makes a significant contribution to knowledge of Late Ordovician biodiversity.
SOMMAIRE
Les lagerstätten de conservation, ces sédiments contenant des fossiles d’organismes à corps mou ou légèrement sclérotisés particulièrement bien conservés, sont très rares dans les strates ordoviciennes. Trois sites d’importance sont connus dans des roches de l’Ordovicien supérieur à Cat Head, Manitoba, soit McBeth Point, William Lake et Airport Cove. Ces sites sont situés dans deux bassins sédimentaires distincts: les deux premiers sont situés dans le bassin de Williston, tandis que le second est situé dans le bassin de la baie d’Hudson. Les trois sites contiennent des fossiles marins, mais chacun présente un assemblage différent, chacun montrant une composante distincte de la diversité biologique d’alors. Les groupes les plus importants représentés, dans un ou plusieurs de ces sites, sont les algues, les éponges, les cnidarian medusae (méduses), les conularides, les trilobites, les euryptérides, xiphosurides (limules) et pycnogonides. Les différents biotopes reflètent les conditions de dépôt de chaque site. Nombre de ces fossiles sont inconnus ailleurs dans l’Ordovicien, au niveau de la famille ou du taxon supérieur de la classification. Ainsi, la province du Manitoba offre-telle une contribution importante à la connaissance de la biodiversité de l’Ordovicien supérieur.
INTRODUCTION
1 Konservat-Lagerstätten (sing. Lagerstätte) are deposits that preserve entirely soft-bodied or lightly sclerotized organisms, or the soft anatomy of forms known otherwise only from their mineralized or decay-resistant anatomy. Such deposits are unusual in the fossil record, because unmineralized tissues are typically broken down entirely by postmortem scavenging and decomposition. Konservat-Lagerstätten are of scientific importance both in establishing the evolutionary histories of organisms that would be otherwise unknown as fossils, and in providing exceptional evidence of long-vanished biological communities.
2 Life in the world’s oceans increased in diversity through much of the Ordovician Period (488 to 444 million years ago), to the extent that we now talk about the Great Ordovician Biodiversification Event, or GOBE (Webby et al. 2004). This event, of unprecedented scale, was comparable in significance to the famous Cambrian Explosion of about 540 million years ago (Webby 2004). Although the great majority of our knowledge of the Ordovician event is based on the record of fossils with preservable hard parts, important evidence about Ordovician biodiversity may also be found in Konservat-Lagerstätten.
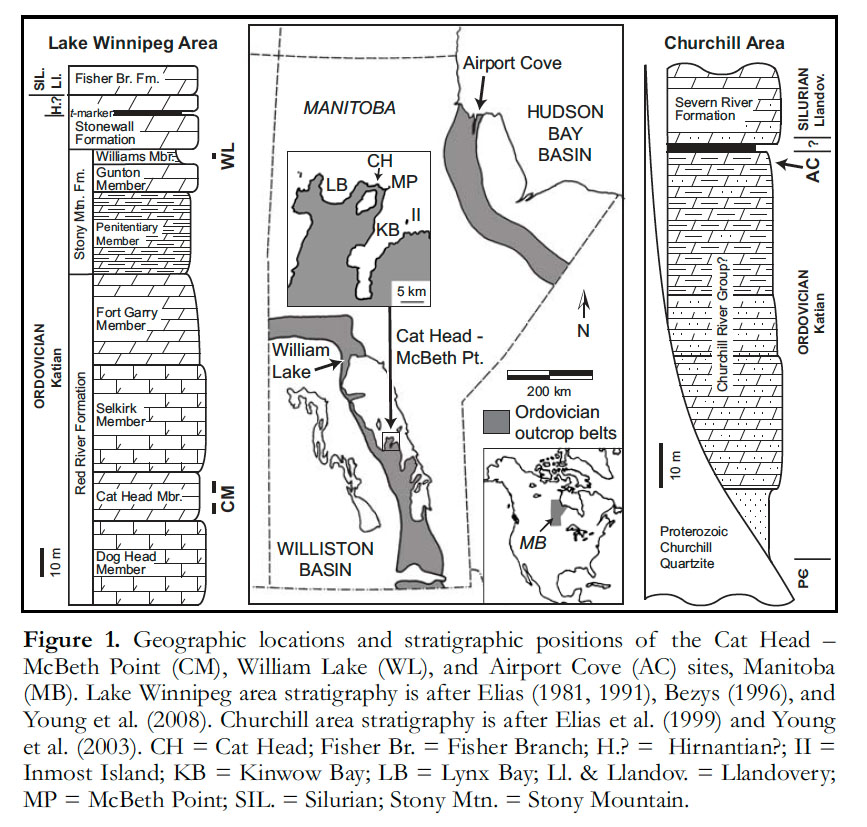
3 Evidence of this biodiversity is, however, tantalizingly scarce, as Ordovician Konservat-Lagerstätten are rare globally. Soft tissue preservation for this time period is far less well documented than for the preceding Cambrian or, indeed, for the subsequent Silurian. Manitoba is fortunate to be endowed with three distinct sites or areas known to contain significant Ordovician soft tissue preservation. These are at Cat Head (or the Cat Head – McBeth Point area), Airport Cove, and William Lake (Fig. 1). Manitoba is still little-explored geologically in comparison with many other parts of the world, and there are probably additional sites waiting to be found. This province may thus make a globally significant contribution to our understanding of Late Ordovician bio-diversity. The three known sites have not yet been thoroughly documented, and much still remains to be learned; studies are ongoing.
 Display large image of Figure 1
Display large image of Figure 14 Ordovician rocks in Manitoba were deposited in two distinct sedimentary basins: the Williston Basin to the southwest and the Hudson Bay Basin to the northeast (Elias 1991; Young et al. 2008; Fig. 1). Of the three Lagerstätten, the Cat Head and William Lake biotas occur in the Williston Basin, while the Airport Cove biota is in the Hudson Bay Basin. The Cat Head biota, of mid Katian (‘early Maysvillian’) age, is significantly older than the other two, which are considered to be of latest Katian (‘late Richmondian’) age (Fig. 1).
5 Ordovician rocks in Manitoba record several cycles of sea level rise and fall within the interior of the paleo-continent called Laurentia, which makes up most of modern North America (Elias 1991). Each of the three Lagerstätten represents conditions in a particular part of a recognized sea level cycle. Cat Head strata (Fig. 2a) are generally considered to represent deeper water, quiet conditions near the transgressive maximum of the Red River Cycle. Strata at William Lake (Fig. 2b, c) record very restricted shallow marginal marine conditions near the regressive extreme at the top of the Stony Mountain Cycle, while the Airport Cove rocks (Fig. 2d-f) were also deposited under restricted marginal marine conditions, possibly during an interval of sea level fluctuation near the extreme of a regression. As a result of the distinctions in age and paleoenvironment, the Cat Head biota, dominated by groups such as trilobites and non-calcareous algae, and distinguished by an extraordinarily diverse conulariid fauna (six species), is quite different from the horseshoe crabs and eurypterids that characterize the other two Lagerstätten.
 Display large image of Figure 2
Display large image of Figure 26 On a global level, Ordovician soft-bodied fossils are very rare. Perhaps the two most remarkable assemblages are in Gondwanan Africa: in the Lower Ordovician Fezouata formations of Morocco, and the Upper Ordovician Soom Shale of South Africa. The Fezouata biota, which seems to exhibit ‘Burgess Shale-type’ preservation, constitutes an extraordinary array of sponges, annelid worms, and a great variety of superbly-preserved arthropods, along with graptolites, conulariids, hyolithids and other molluscs, brachiopods, bryozoans, and echinoderms (Van Roy 2006; Vinther et al. 2008; Van Roy et al. 2010). These fossils apparently represent creatures that lived on an open marine shelf. The Soom Shale assemblage, deposited in cold temperate shelf conditions, comprises a more modest assortment of fossils, including rare naraoiid arthropods, eurypterids, straight-shelled ‘nautiloid’ cephalopods, conodont animals, and anaspid-like ‘fishes’ (Aldridge et al. 2001; Selden and Nudds 2004; Aldridge, pers. comm. 2004). More recently, work on Middle and Late Ordovician sequences in Avalonian Wales has revealed sites preserving mixed shelly and soft-bodied biotas in a variety of preservational modes (Botting et al. 2011; Botting and Muir 2012).
7 In other parts of Laurentia, many of the known Ordovician Lagerstätten represent marginal marine environments. The Middle Ordovician Winneshiek Lagerstätte of Iowa (Liu et al. 2006, 2011; Witzke et al. 2011) is quite diverse, including conodonts, primitive fishes, lingulid brachiopods, phyllocarid crustaceans, and eurypterids. The Middle Ordovician rocks at Douglas Dam, Tennessee (Caster and Brooks 1956; Dunlop et al. 2004) and the uppermost Ordovician Georgian Bay Formation of Manitoulin Island, Ontario (Stott et al. 2005) hold low-diversity assemblages of mostly chelicerate arthropods. Body fossils in the Upper Ordovician Harding Sandstone of Colorado are diverse but poorly preserved (Allulee and Holland 2005), and additional undescribed marginal marine fossils are coming to light in places such as Wisconsin. In a much deeper water deposit, spectacularly preserved trilobites occur in the famous Upper Ordovician Beecher’s Trilobite Bed of New York State (Briggs et al. 1991; Etter 2002; Selden and Nudds 2004).
8 The Manitoba sites are providing data that are not duplicated by or subsidiary to that from any other place. Rather, every soft tissue site discovered to date is contributing a separate, complementary piece of the puzzle that is Ordovician biodiversity.
CAT HEAD – McBETH POINT
9 In the Cat Head – McBeth Point area of Lake Winnipeg’s North Basin, the Cat Head Member of the Red River Formation contains a considerable variety of fossils, combining soft-tissue preservation of certain groups with other forms representing a normal marine mineralized biota. It is most famous for its remarkable assemblages of marine algae (seaweeds), but there are also conulariids, trilobites, lingulid brachiopods, and many other groups. The fossils are found along the west shore of Lake Winnipeg between McBeth Point, Cat Head, and Lynx Point, north and west of Kinwow Bay (Fig. 1). There, about 12 m of strata are exposed in cliffs along the lakeshore, over a horizontal distance of about five km (Fig. 2a). In addition, similar fossils are found at Inmost Island in the middle of Kinwow Bay (formerly known as Birch Island or Kinwow Bay Island); these are probably derived from slightly older beds. All of the fossiliferous units are considered to belong to the lower part of the Cat Head Member of the Red River Formation, of mid Katian age (Fig. 1).
10 Cat Head and McBeth Point are prominent topographic features on the North Basin, and the latter forms an important anchorage. Thus, it is hardly surprising that these spectacular fossils were known from the early days of collecting in this region. Fossils in the Cat Head area were apparently first systematically collected by members of Geological Survey of Canada (GSC) field parties, including T.C. Weston in the 1880s and D.B. Dowling and L.M. Lambe in 1890s (Fig. 2a inset). Further collecting for the GSC was done by G.W. Sinclair and others in the 1950s. These sites were visited in 1997 by staff from The Manitoba Museum and associates from the Manitoba Geological Survey and Royal Ontario Museum, and again by The Manitoba Museum in 2006. Many other specimens now in collections have been picked up by fishermen and pleasure boaters over the years. The first description of some of the fossils from this area was by Whiteaves (1897); shortly thereafter, Dowling (1900) produced a very good description of the basic geology and included a substantial listing of the fossils. Sporadic paleontological studies since that time have most notably included the description of palynomorphs, cephalopods, and sponges in a collaborative GSC Bulletin (McGregor and Cramer 1971; Flower 1971; Rigby 1971) and a study of the remarkable non-calcareous algae (Fry 1983). Major collections of Cat Head fossils are located at The Manitoba Museum, the University of Manitoba, and the Geological Survey of Canada (Ottawa).
11 The unusual fossils in the Cat Head – McBeth Point area are derived from bedded fine-grained dolostones that lack storm deposits and extensive burrow mottling. Chert nodules of variable colour and form are abundant in some beds. The lithology, and the abundance of fossils interpreted as algae, are consistent with deposition in the photic zone, but probably below storm wave base. This interpretation fits with inferred transgressive-regressive cycles for the Williston Basin, which place the Cat Head biota in the deepest water conditions of the Red River Cycle (Elias 1991). It has, however, also been suggested that the conditions were those of a restricted marine environment having little contact with the open sea (McGregor and Cramer 1971).
12 Fossils, generally rare but abundant at some levels, include macroscopic non-calcareous algae (Figs. 2a inset, 3a, 3b), possible hydro-zoans (Fig. 3c), palynomorphs, sponges (Fig. 3d), , conulariids (Fig. 3e, 3f), ‘nautiloid’ cephalopods, gastropods, articulated trilobites (notably Isotelus sp.; Fig. 3g), brachiopods (obolids, lingulids, orthids, and strophomenids; Fig. 3h), , crinoids, dendroid graptolites (Fig. 3i), and the problematic Sphenothallus (Whiteaves 1897; McGregor and Cramer 1971; Flower 1971; Rigby 1971; Fry 1983; Westrop and Ludvigsen 1983; Jin and Zhan 2001; Robson and Young in press).
 Display large image of Figure 3
Display large image of Figure 3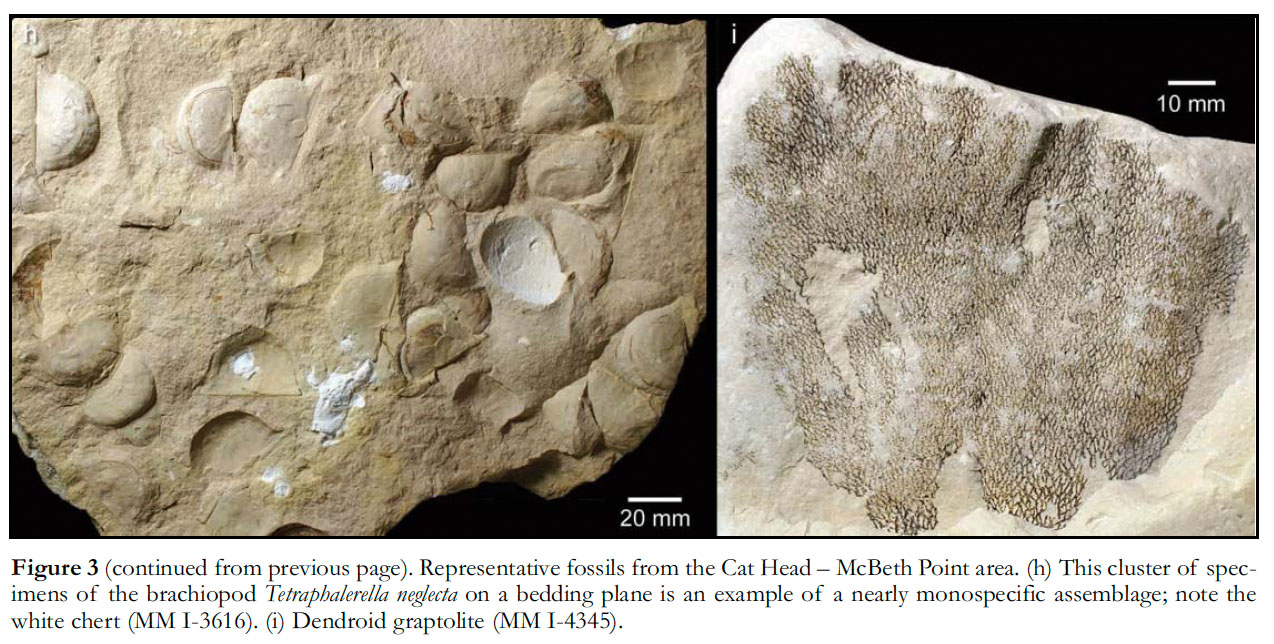 Display large image of Figure 3
Display large image of Figure 313 The soft algae (seaweeds) are among the best to be found anywhere in Early Paleozoic rocks, and have been interpreted as rhodophytes, heterokontophytes, and chlorophytes (Fry 1983). The conulariids and trilobites are remarkable for their articulation, indicating that they were buried under low-energy conditions. The beds within which fossils are abundant typically contain one or very few species, suggesting that briefly favourable conditions resulted in ‘blooms’ of life (Fig. 3h).
14 Preservation of the Cat Head fossils has not been studied in detail. Many of those with mineralized calcareous skeletons, such as trilobites and strophomenid brachiopods, appear to be preserved through replacement by fine-grained dolomite and chert. Phosphatic or chitinophosphatic skeletons, such as those of conulariids and lingulid brachiopods, are wonderfully preserved and have the appearance of unmodified material. The seaweeds occur as films that include carbon and iron oxide (which may have originally been pyrite). The preservation of the seaweeds is such that they are sensitive to light and weather. If left exposed outside, they fade and become more ‘rusty’ as carbon is lost. Hydroids and dendroid graptolites are preserved as carbon, apparently with some pyrite. A few rare fossils, such as sponges, are occasionally found as pyritic replacements of soft tissue within chert nodules.
15 The overall composition of the Cat Head biota is similar to those of certain younger, Silurian Lagerstätten from elsewhere in Laurentia. These latter, termed the DG biotas by Kluessendorf (1994), are characterized by dendroid graptolites associated with organisms such as algae, trilobites, and sponges. The DG biotas were preserved under a variety of conditions, but appear to be generally linked by rapid burial in places where anoxic bottom waters were associated with topography. Like the Cat Head assemblages, many of the constituents represent normal-marine organisms preserved under unusual circumstances.
WILLIAM LAKE AND AIRPORT COVE: SHARED CHARACTERS AND SHARED DISCOVERY
16 The William Lake and Airport Cove sites represent distinct depositional environments, but both were formed in restricted and marginal marine settings, in very shallow water and shoreline to nearshore conditions. Both are of latest Katian age, and they share a number of biotic elements, though with differing relative abundances: most notable are xiphosurids (horseshoe crabs) and eurypterids, but there are also non-calcareous algae, conodonts, lingulid brachiopods, gastropods, and large phosphatic or chitinophosphatic tubes of uncertain affinity (Young et al. 2007). These assemblages are strikingly different from normal marine biotas of nearby units. They completely lack corals, trilobites, and echinoderms, while rhynchonelliformean brachiopods and ‘nautiloid’ cephalopods are rare.
17 Fossil sites such as those at William Lake (Fig. 4) and Airport Cove (Fig. 5) are poorly represented in the geological record because they form in shoreline environments that migrate, are limited in area, and have low preservation potential for body fossils. Both biotas occur in what initially appeared to be monotonous and unfossiliferous dolomudstones of generally similar character, and these units were largely ignored until the unusual fossils were discovered. It is, in fact, the similarities between the biotas and lithotypes that resulted in the discovery of both sites within the past dozen years.
 Display large image of Figure 4
Display large image of Figure 4 Display large image of Figure 5
Display large image of Figure 518 Intensive study of these sites arose from two fortuitous discoveries of arthropod fossils. In the 1990s, J.-D. Fournier, a geologist working in the Grand Rapids Uplands, spent his spare time searching for flat slabs of rock to be used as substrates for artwork. He discovered an area where suitable slabs had been piled up, and selected appropriate pieces on which to paint. Later, noticing fossils on the reverse sides of slabs, he brought these to the attention of Manitoba Survey geologist Ruth Bezys, who passed the information on to one of us (G.Y.; see Fig. 4e). Manitoba Museum staff first visited the area briefly in 2001, finding an articulated eurypterid and other fossils, but the bedrock source for this material could not be located until a much longer visit in the autumn of 2004. This locality, now called the William Lake site, has subsequently been visited many times, and since 2007 a systematic, bed-by-bed collection has been carried out.
19 In 2000, then-student Sasha Herwig was assisting Robert Elias and Graham Young with paleoecological field research approximately 650 km northeast of William Lake, at the cove north of Churchill Airport. Examining loose surface material, she discovered the beautiful anterior shield of a small eurypterid-like arthropod and associated fragments (Fig. 5g). However, as at William Lake, the bedrock source could not initially be located.
20 At William Lake in June, 2005, three of us (E.D., S.R. and G.Y.) collected several unusual fossils, including the first articulated specimen of a creature that was to be later described as the early horseshoe crab Lunataspis aurora (Rudkin et al. 2008). Examining the rocks in detail, we realized that similar bedrock could be found in some areas of Airport Cove. At the latter site a few weeks later, a substantial area of loose angular blocks of dolostone having the appropriate appearance was located; splitting these blocks, within 30 minutes additional specimens of Lunataspis were discovered, along with eurypterids and a variety of other fossils! The Airport Cove site was revisited in July, 2011; it now appears certain that it consists of bedrock that had been largely frost-wedged in situ (i.e., very little transport had occurred), but there is no exposed outcrop. All visible fossils at that site have now been collected.
21 All xiphosurids from both sites have, to date, been assigned to Lunataspis aurora, which is among the oldest known horseshoe crabs, representing a lineage still found in marginal marine environments today (Rudkin et al. 2008; Rudkin and Young 2009; Figs. 4i, j; 5h, i). Currently, it is difficult to determine how many eurypterid taxa are present among the two biotas because the superb material shows different modes of preservation and a number of unusual features. These important fossils, particularly those from William Lake, form a significant addition to the sparse Ordovician record of the Eurypterida (Cuggy et al. 2010; Figs. 4e-g, 5e, f).
WILLIAM LAKE SITE: PALEOENVIRONMENT AND PALEOBIOLOGY
22 The William Lake site is notable both for the remarkable fossil groups it shares with Airport Cove, and for those that are unique to it: jellyfish and other gelatinous zooplankton, and bizarre arthropods such as pycnogonids (‘sea spiders’). This site is located on the Grand Rapids Uplands between Grand Rapids and Ponton, in an area rich in fossiliferous Ordovician and Silurian bedrock outcrops. The William Lake fossils occur in the Williams Member of the Stony Mountain Formation (Fig. 1); the stratigraphic position and associated conodonts (Rhipidognathus symmetricus, Drepanoistodus? sp., and Aphelognathus sp.) are consistent with a Late Ordovician (late Katian) age (Young et al. 2007).
23 Thin-bedded dolomudstones of the Williams Member represent restricted, shallow, and very low-energy conditions in a marginal marine environment (Fig. 2b). The conodont assemblage in this interval (Rhipidognathus biofacies) indicates the shallowest conditions for the Late Ordovician of the Williston Basin (Nowlan and Haidl 2001). The fossiliferous beds occur within a shallowing-upward succession, above burrow-mottled rocks bearing normal-marine fossils, and below laminated, virtually unfossiliferous dolomudstones in which some bedding planes are covered with salt crystal lattices.
24 Over the past several years, detailed collecting has sampled an interval about 2.5 m thick; most Lagerstätte-type fossils occur within the middle 1.5 m of this interval. Some bed surfaces are broadly rippled or hummocky. Cut and polished vertical sections reveal parallel lamination and trough cross-lamination; channel-like features cut across the laminae in places. Some beds consist of monotonous mudstones, while others are replete with mud rip-up clasts. Features such as apparent dewatering structures and microbial mat textures are occasionally visible. The fossils are generally rare, although narrow intervals locally contain common to abundant lingulid valves, sometimes mixed with arthropod fragments and other shelly material (Fig. 2c). These horizons commonly coincide with bed partings, whereas beds between these fossil-rich horizons show less distinct internal structure, and local lenses or layers of monotonous dolomudstone.
25 The distribution of fossils indicates variable environmental conditions that were sometimes unfavourable for all fossil groups, but occasionally favourable for particular groups. Some groups occur through much of the studied interval, although they are rare in most beds, e.g. lingulid brachiopods, eurypterids (Fig. 4e-g), gastropods, and the horseshoe crab Lunataspis aurora (Fig. 4i, j). Other fossils are narrowly concentrated in specific beds. Jellyfish, which include hydrozoan and possibly scyphozoan medusae, are common or abundant at some horizons in the lower to middle part of the interval (Fig. 4a, b) but nearly absent from the upper part. The large tubes of undetermined affinity also occur only in the lower part (Fig. 4d). Probable algal fronds are common only in a single upper bed, a thinly laminated unit that is unusual for the occurrence of ostracodes and tiny juvenile specimens of Lunataspis (Fig. 4j).
26 Some of the strangest fossils at William Lake are represented by a handful of examples. Several pycnogonid specimens are the geologically earliest known adult sea spiders, and the first report of this group in the North American fossil record (Rudkin et al. in press; Fig. 4h). Remarkable fossils that probably represent the comb rows of numerous ctenophores (comb jellies) occur in a cluster within a single slab (Fig. 4c).
27 The fossils at William Lake are variably preserved depending on taxonomic group, and on the type of stratum in which they occur. Those showing the finest preservation, the medusae (jellyfish) and articulated arthropods, are found in largely homogeneous layers or lenses of rapidly deposited mudstone (Fig. 2c). The medusae are preserved as sparry dolomite that is slightly enriched in iron and silica, and they are surrounded by halos of degraded pyrite.
28 The original quality of preservation of some of the arthropod fossils may have been much higher, but it is likely that they lost easily weathered organic components as the host beds were exposed at or near the surface for at least the past several thousand years (their upper surfaces are glacially striated). The best-preserved eurypterids are largely mouldic but include some organic material and pyrite (Fig. 4g). Xiphosurids and pycnogonids at this site, on the other hand, show no evidence of organic skeletal material; these differences are probably related to differing original exoskeletal structures and compositions.
29 Lingulid brachiopods, gastropods, and disarticulated arthropods are most common in thin, distinct horizons that also show evidence of horizontal burrows (Fig. 2c). These layers may include lags that accumulated during times of reduced sedimentation, as channel fills, and as concentrations in the troughs of broad ripples. The fossils within them are generally preserved as moulds, although patches of organic material are occasionally found. Almost all lingulids occur on these sorts of bedding plane surfaces, suggesting that they were not buried in life position.
30 All evidence suggests that the William Lake fossils were preserved in an intermittently harsh, unpleasant place, possibly a ponded intertidal mudflat, or a restricted subtidal lagoon (cf. Pratt 2010; Rankey and Berkeley 2012). The climate was hot, there was little wave or current motion, and the water was very shallow and at times hypersaline. The sediment was an anoxic or dysoxic fine soupy mud. It is possible that this was not generally a place where creatures permanently lived; rather, it was where they languished and died. This seems to be the case for the cnidarian medusae. Horizons containing abundant medusae may record jellyfish blooms that had drifted into unfavourable peri-tidal environments. The ‘halos’ around many of the jellyfish may have resulted from anaerobic conditions associated with decomposition. Small horizontal burrows in the substrate very rarely intersect the jellyfish; rather, they tend to stop outside a zone of visible disseminated iron oxide that surrounds each medusa. Modern blooms can be sufficiently profuse that decomposing jellyfish cause dramatic temporary declines in oxygen content of lagoonal sediments (West et al. 2009); some beds at William Lake may provide possible evidence of this phenomenon in the distant past.
31 Most of the more ‘normal marine’ forms at William Lake, such as ‘nautiloid’ cephalopods and rhynchonelliformean brachiopods, possess shelly hard parts that would have been readily shifted or floated by very slight wave or current action. It is likely that some of them were transported from environments that had more favourable conditions, while others may have had specific adaptations that permitted them to live in this unusual environment (Fig. 4d).
32 Many of the eurypterids and horseshoe crabs at this site are small and may represent juveniles (Fig. 4j). These might have blundered or been washed into conditions where they could not survive, or it is possible that the very shallow water served as a ‘nursery’ from which they would have migrated into more open and unprotected waters as they grew. A lagoonal nursery situation has been suggested for eurypterids elsewhere (see Braddy 2001). Interestingly, the majority of articulated eurypterid remains exhibit orientations suggesting they represent moulted individuals, rather than intact carcasses. It is also notable that we are beginning to see significantly larger eurypterids as we collect lower in the section, into rocks that show evidence of having been deposited under less harsh conditions.
33 The composition of the William Lake biota is similar to those of the younger ‘EP biotas’ that occur at Upper Silurian sites in central Laurentia (Kluessendorf 1994). EP biotas are characterized by eurypterids and other arthropods (sometimes including xiphosurans), often associated with hypersaline conditions. Like the William Lake biota, the EP organisms were preserved in anoxic conditions in peritidal paleoenvironments.
BIOTA AND LITHOLOGY AT THE AIRPORT COVE SITE
34 The fossil assemblage at Airport Cove differs from that at William Lake in the relative abundance of beautiful seaweeds, particularly non-calcifying dasycladacean green algae, the occurrence of polychaete worms in the form of articulated scolecodont assemblages, and the presence of organically-preserved arthropod cuticle (Young et al. 2007). Among the shared constituents, eurypterids are notably rarer here than at William Lake, and they are more frequently disarticulated. As is the case at William Lake, the fossils at Airport Cove are generally sparsely distributed and difficult to find, but locally concentrated.
35 Airport Cove is located a few kilometres east of the Town of Churchill, and immediately north of Churchill Airport (Fig. 1). Ordovician and Silurian sedimentary rocks in this area surround a remarkable paleo-archipelago composed of Proterozoic Churchill quartzite (Nelson and Johnson 2002; Young et al. 2003). Elsewhere in the Hudson Bay Basin, several distinctive Upper Ordovician lithostratigraphic units are recognized (Nelson 1963, 1964; Norford 1971; Norris 1993). In the Churchill area, however, the distinctions between units are complicated by the archipelago’s influence on local sedimentation (Elias et al. 1999).
36 The fossils occur in angular, relatively fresh slabs found loose within a narrowly-defined area in Airport Cove (Fig. 2d). These slabs of distinctive greenish dolostone have yielded conodonts consistent with a Late Ordovician (Late Katian) age: Rhipidognathus symmetricus, ?Aphelognathus divergens, and Drepanoistodus suberectus (Young et al. 2007). Nearly identical rocks occur near the top of a drillcore extracted nearby within the cove, in a unit that was previously tentatively assigned to the Churchill River Group (Young et al. 2003; Young et al. 2007). Recent analysis of the upper portion of the 100 m of section observed in drillcore from Airport Cove suggests that these dolostone beds occur within an interval that may instead belong to the Port Nelson Formation (Wheadon 2011).
37 The lithotype in which the Airport Cove fossils are found is a dolomudstone with distinctive grey-green to buff laminae 2-4 mm thick, alternating with thinner dark grey-green pyrite-rich bands (Fig. 2e). In places, polished vertical surfaces show repeated small-scale lenticular to wavy bedding characteristic of tidal environments (Davis 2012). Sedimentary features include apparent microbial mats, soft sediment deformation, and evaporite crystal moulds. These features, together with the unusual macrofossils and associated conodonts, all indicate very shallow and restricted conditions. Since the site is almost adjacent to an angular unconformity against the Proterozoic quartzites that define the cove, this was clearly a nearshore to shoreline environment, beside a quartzite paleoisland (Fig. 2d, f).
38 Well-preserved macrofossils tend to occur within the darker bands, while scolecodonts and small calcareous fossils are locally common in the paler intervals. Macrofossils are preserved as pyrite, as carbonate, and as organic matter (cf. Briggs 2003). The best preserved fossils at Airport Cove include arthropods and algae, both of which are largely preserved as organic material. Arthropod sclerites are often disarticulated and broken, but cuticular detail is remarkable (Fig. 5e). Specimens of the horseshoe crab Lunataspis aurora exhibit a combination of organic material, localized pyrite, and mouldic preservation (Fig. 5h, i). Dasycladalean algae show very finely-detailed carbonaceous compression preservation (Fig. 5a, b). Scolecodonts are common on bedding planes, occasionally as articulated apparatuses (Fig. 5c).
39 As is the case for William Lake, the Airport Cove biota is somewhat similar to the younger ‘EP biotas’ that occur at Upper Silurian sites in central Laurentia (Kluessendorf 1994), though it appears that water circulation was at times more open than that at William Lake (i.e. it may have been physically less removed from open marine conditions). The mixture of articulated and disarticulated fossils at Airport Cove may indicate variation through time in the amount of wave and current activity. The alternation of laminae in the host dolostone is consistent with this: the darker laminae may represent intervals of stagnation, or possibly record the development of microbial mats or crusts.
SIGNIFICANCE
40 Many of the fossils recovered at the three exceptional Manitoba sites are quite different from those found in other Late Ordovician settings, within Laurentia and globally. They record the presence of rare or otherwise unknown taxonomic groups and thus contribute key data points to our understanding of rapidly expanding marine biodiversity associated with the GOBE. They also provide critical insights into the evolutionary histories of groups of organisms with low preservation potential. The authors are most actively studying the William Lake biota, so the most detailed understanding pertains primarily to that site, which represents deposition in extremely restricted and shallow-water conditions. The other sites, though less well understood, supply complementary information on different settings.
41 Together, William Lake and Airport Cove provide unique data about tropical shoreline communities prior to the Late Ordovician extinctions. Almost half the taxa (ordinal level or higher) are rare or unknown elsewhere in the Ordovician of North America. Considering taxonomic groups, 33% of fossil groups at William Lake are calcareous, in contrast with normal marine carbonates lower in the Stony Mountain Formation, where 95% are calcareous (Young et al. 2010). In total numbers, fossils with calcareous skeletons are remarkably rare at William Lake and Airport Cove, making up only four to five percent of the total numbers of macrofossils at each.
42 Many of the fossils found at William Lake and Airport Cove belong to still-extant groups such as horseshoe crabs and lingulid brachiopods. As we consider these biotas, it is easy to fall into the trap of thinking of them as though they represent environments very similar to those found in modern shoreline conditions. We must not, however, lose sight of the fact that the world in which these organisms lived was very different, with a higher global mean surface temperature, lower atmospheric oxygen, extremely high atmospheric CO2 (possibly up to 15x modern pre-industrial levels), considerable volcanic activity, and global sea levels up to 220 m above modern levels (Trotter et al. 2008; Servais et al. 2009)! The occurrence of these very long-lived groups is not evidence of unchanging conditions. Rather, it demonstrates the considerable adaptability and resilience of these organisms.
43 All three sites include many forms that are not preserved in standard normal marine Late Ordovician assemblages, and each has the potential to improve our understanding of bio-diversity during this critical interval in Earth’s history.
Funding for this research was provided by NSERC, The Manitoba Museum Foundation, and the Royal Ontario Museum Governors. We are grateful to Norman Aime, Peter Fenton, Marion Foster, Brian Iwama, Christine Kaszycki, Lori Stewart, Marjorie Turton, and David Wright, for their participation in fieldwork at one or more of these fossil sites. We would like to thank Ruth Bezys, J.-D. Fournier, and Sasha Herwig for their role in the initial discovery of the William Lake and Airport Cove sites. We have appreciated discussions of these rocks and fossils with many individuals, including Nicholas Butterfield, Robert Elias, and Whitey Hagadorn. David Greenwood is thanked for providing kind access to microscope facilities at Brandon University, and for discussions. Our research has benefited from Godfrey Nowlan’s conodont analyses, and from collaboration with the Manitoba Geological Survey and its drill crew (Doug Berk, Gerry Benger, and Vio Varga), both of which contributed to the stratigraphic and pale-oenvironmental understanding of the William Lake and Airport Cove sites. This manuscript was improved considerably thanks to thorough reviews by Jakob Vinther and Joe Botting.