Series
Great Canadian Lagerstätten 1.
Early Eocene Lagerstätten of the Okanagan Highlands (British Columbia and Washington State)
SUMMARY
The Early Eocene Okanagan High lands series of lacustrine shale and coal deposits, in far western North Ameri ca, constitutes a significant group of fossil sites with exceptional preserva tion of a diverse suite of organisms (Lagerstätten). With contemporaneous basins arrayed across about 1000 kilo metres of southern British Columbia and northern Washington, these sites offer a unique opportunity to examine the paleoecology of terrestrial commu nities spanning a temperate, low-sea sonality landscape in a montane setting during a time of generally warm tem peratures across the globe. The Okana gan Highlands sites provide an unpar alleled comparative framework within which to examine this major turning point in terrestrial community develop ment during the emergence of their broad modern character.
SOMMAIRE
La série de dépôts lacustres de schiste et de charbon du début de l’Éocène des hautes terres d’Okanagan, aux con fins de l’ouest de l’Amérique du Nord, constituent un groupe important de sites fossiles particulièrement bien con servés de suites d’organismes diverses (Lagerstätten). De nos jours, ces sites forment en une bande d’environ 1 000 kilomètres, depuis le sud de la Colom bie-Britannique jusqu’au nord de l’État de Washington. Dans le contexte de réchauffement climatique, c’est l’occa sion ou jamais d’étudier la paléoécolo gie de communautés terrestres dans des conditions climatiques modérées dans un paysage de montagne à faible saisonnalité. Les sites des hautes terres d’Okanagan représentent un cadre de comparaison sans pareil permettant d’étudier les effets de ce tournant majeur sur le développement des prin cipales caractéristiques modernes de la communauté terrestre.
INTRODUCTION
1 Lagerstätten (Konservat-Lagerstätten in particular) are fossil assemblages that combine fine preservation of features not typically fossilized (e.g., soft tissues, hairs, colour patterns) with high biological diversity, providing exceptional records of past life (Seilacher 1970). Famous examples include the Eocene sites at Florissant, Colorado (Meyer 2003) and Messel, Germany (Schaal and Ziegler 1992), and the Cambrian Burgess Shale of British Columbia (Gould 1989). A series of Early Eocene sites collectively called the Okanagan Highlands also fits this description well. These occur across an approximately 1000 km transect, stretching from northern Washington State to central British Columbia (Fig. 1), in depositional basins where lacustrine shales and coal deposits fill grabens associated with uplift and regional volcanism (Ewing 1980). The deposits reveal ancient forest, lake and swamp life in temperate upland communities within the context of a warm and seasonally equable world, including a rich suite of plants, insects, fish, feathers, and (rarely) bird and mammal skeletal remains (Figs. 2, 3), as well as microfossils such as diatoms and pollen. Megafossils are commonly spectacularly well preserved as compressions featuring colour patterns on insects’ wings, exquisite whole flowers of Florissantia, and delicate ferns like the floating Azolla (Fig. 4), and showing minute details of epidermal cell patterns of leaves (Fig. 5) and membrane hairs on the wings of tiny fungus gnats (Fig. 6). These fossils reveal communities in their environmental contexts some fifteen million years after the end-Cretaceous mass extinction event, affording an unprecedented opportunity to examine the origin and development of large-scale patterns of diversity, biogeography, evolution and climate.
 Display large image of Figure 1
Display large image of Figure 1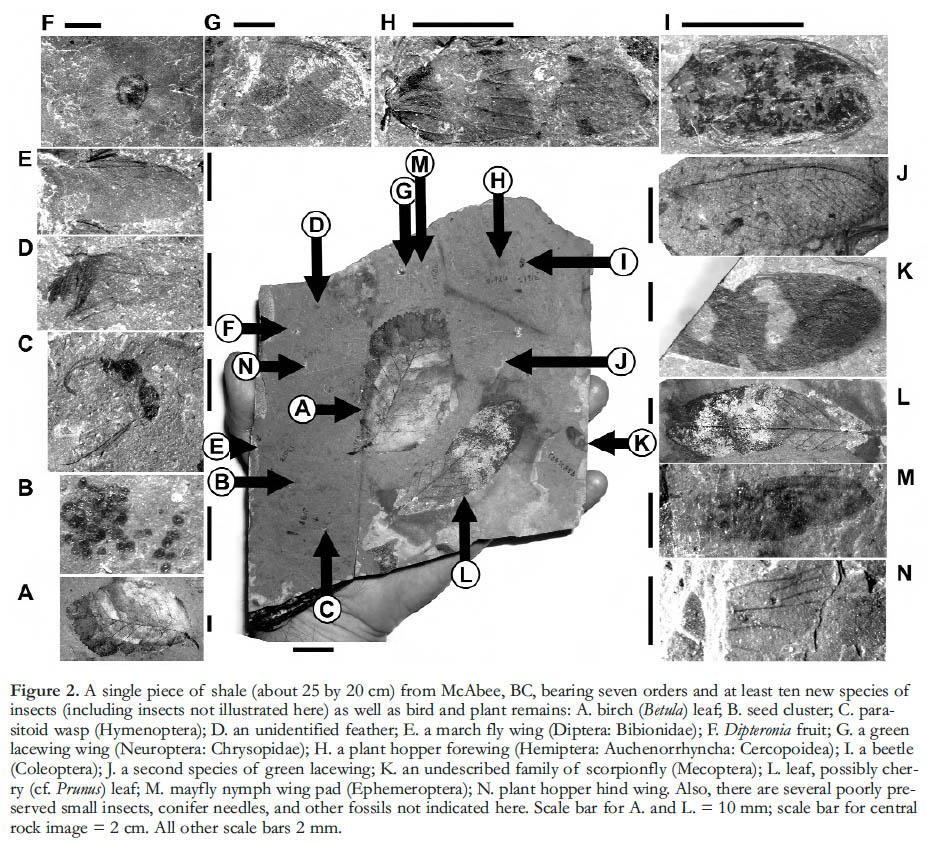 Display large image of Figure 2
Display large image of Figure 2 Display large image of Figure 3
Display large image of Figure 3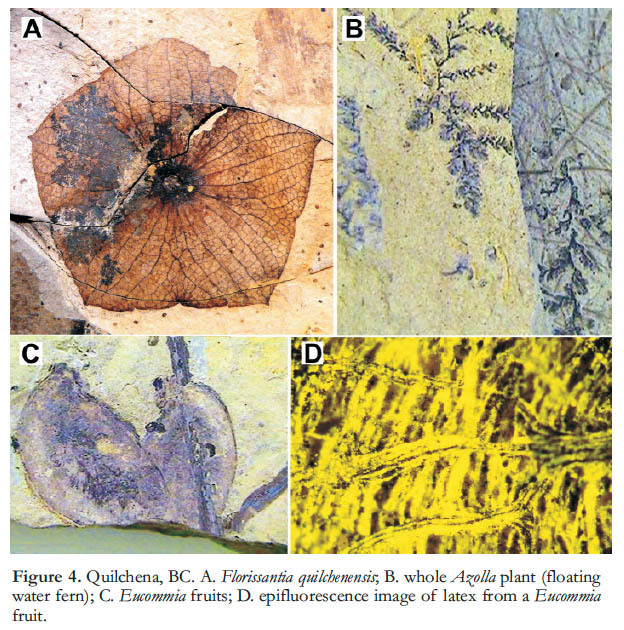 Display large image of Figure 4
Display large image of Figure 4 Display large image of Figure 5
Display large image of Figure 5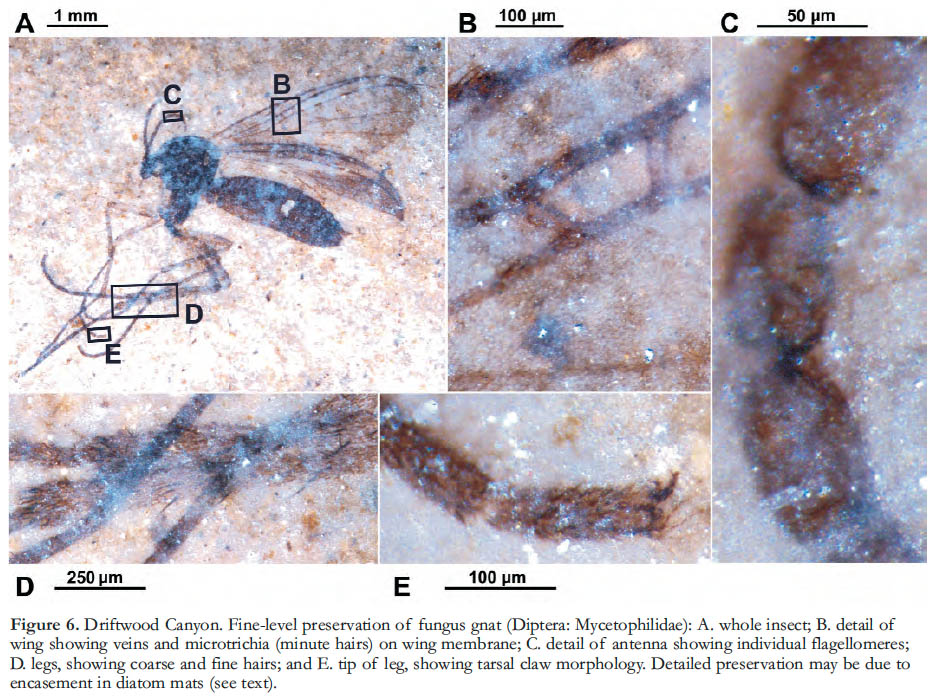 Display large image of Figure 6
Display large image of Figure 6HISTORY OF WORK
2 Until the latter part of the 20th century, the remoteness of the Okanagan Highlands hampered its scientific study compared with more accessible contemporaneous localities, such as those near railway lines in mid-continental USA (e.g. Meyer 2003). When British Columbia entered Confederation in 1871, following a series of gold rushes, the federal government sent geologists to assess its coal and mineral wealth. Among them was George Mercer Dawson (1879, 1895), who reported plant and insect fossils in sedimentary rocks of the Interior, later called the Okanagan Highlands (earliest use apparently by Wehr and Schorn 1992). These early collections resulted in a variety of studies in paleoentomology (Scudder 1877; Handlirsch 1910); paleobotany (e.g. J.W. Dawson 1879, 1883, 1891; Penhallow 1908; Berry 1926, 1929; Brown 1935, 1937, 1939, 1940); and rare mammals (two teeth; Russell 1935; Gazin 1953).
3 After this initial burst of activity, only sporadic studies were published until a renaissance in the latter half of the 20th century, beginning with work on pollen in the 1960s (Rouse and Mathews 1961; Mathews 1964; Hills 1965; Hills and Baadsgaard 1967), then gaining momentum in the 70s with the first description of the McAbee flora (Verschoor 1974) and examination of the exquisitely detailed three-dimensional plant fossils of the Princeton chert (Miller 1973; Basinger 1976, 1984; Basinger and Rothwell 1977; Cevallos-Ferriz et al. 1991; Erwin and Stockey 1991; Stockey et al. 1999; Smith and Stockey 2007; and others). The scope of research expanded with work on fish and insects (Wilson 1977a, b, 1978, 1980) and paleobotanical study focusing on the Republic site (e.g. Wolfe and Wehr 1987), leading to a series of papers with a broader regional view, often inspired by talented amateur American paleobotanist Wes Wehr’s promotion of Okanagan Highlands paleontology (e.g. Lewis 1992; Reed 1996; see also Archibald et al. 2005a for Wes Wehr’s obituary). In recent years, interest in Okanagan Highlands fossils has continued to grow (e.g. Basinger et al. 1996; Stockey and Wehr 1996; Wilson 1996a, b; Archibald and Greenwood 2005 and references therein).
PRESERVATION OF FOSSILS
4 Fossils of the Okanagan Highlands are typically preserved in lacustrine shales. Preservation of the stunning detail often seen in these fossils (Figs. 2-6) may be related, at least in part, to their entrapment in diatom blooms and subsequent coating by biofilms, as inferred for Eocene lacustrine-shale Lagerstätten elsewhere (Harding and Chant 2000; Mustoe 2005). Archibald and Makarkin (2006) also suggest that biofilm coating may have lessened taphonomic sorting of insect assemblages by surface area/mass during flotation on the lake surface, thereby increasing fidelity of the fossil assemblages to the original community compositions. Research into lake environments, depositional settings and taphonomic processes in the Okanagan Highlands has resulted in highly influential papers (e.g. Wilson 1980; Barton and Wilson 2005), particularly from work at the Horsefly site, which will be specifically treated by M.V.H. Wilson in a subsequent paper in this Canadian Lagerstätten series.
LIFE AT THE DAWN OF THE RECENT
5 Early workers considered these deposits to be Oligocene or Miocene (Scudder 1878, 1895; G.M. Dawson 1879, 1896). J.W. Dawson (1891) provisionally agreed, but suggested that they might also be Eocene. This was later supported by teeth of the tillodont mammal Trogosus found in the Allenby Formation near Princeton (Russell 1935), then confirmed by K–Ar dating, which indicated Middle Eocene ages throughout the region (Rouse and Mathews 1961). Recent Ar 40 /Ar 39 and U–Pb analyses have pushed back their ages a bit further into the latter half of the Early Eocene (Villeneuve and Mathewes 2005; Moss et al. 2005; Mortensen and Archibald, personal communication).
6 Life in the Early Eocene was making a large step toward modernization following events such as the great extinction at the end of the Cretaceous and the brief, intense interval of global warming at the Paleocene–Eocene boundary (Zachos et al. 2008). Here, in the Okanagan Highlands, numerous groups of animals and plants make their first appearances in the fossil record; for example, genera such as hazel (Corylus) and hornbeam (Carpinus) in the birch family (Betulaceae); the oldest known service-berry (Amelanchier), snow-wreath (Neviusia), and cherry (Prunus) in the rose family (Rosaceae); winter-hazel (Corylopsis) and witch-alder (Fothergilla) in the witchhazel family (Hamamelidaceae); and beech (Fagus) in the beech family (Fagaceae) (e.g. Wolfe and Wehr 1987; Wehr and Hopkins 1994; Manchester and Dillhoff 2004; Radtke et al. 2005; DeVore and Pigg 2007, 2010). We see the oldest records of such insects as seed weevils (Coleoptera: Chrysomelidae: Bruchinae), sweat bees (Hymenoptera: Halictidae), and panorpid scorpionflies (Mecoptera: Panorpidae) (Archibald and Mathewes 2000; Engel and Archibald 2003); and the earliest salmonid fish (Wilson and Li 1999).
7 Mixed with these modern elements were such extinct taxa as the ubiquitous and often beautifully preserved Florissantia (Fig. 4A) and bizarre holcorpid scorpionflies (Archibald 2010), reminding us that this was still a world in transition. These communities were organized in distinctly different manners than we would expect to see today, molded, in large part, by their particular climatic setting.
EARLY EOCENE UPLANDS CLIMATE
8 In the Early Eocene, the world experienced the highest temperatures of the Cenozoic, notably during hyperthermal events, when atmospheric carbon dioxide greatly exceeded modern levels (Zachos et al. 2008; Smith et al. 2010). This was the acme of the ‘greenhouse world’ global climatic regime of warm temperatures and mild winters before the climatic downturn initiating our current ‘icehouse world’ after the close of the Eocene (Zachos et al. 2008).
9 Multiple geologic and paleontological indicators suggest that this region was a temperate upland in this warm world (e.g. Greenwood et al. 2005; Smith et al. 2009; and references therein), with upper microthermal to lower mesothermal mean annual temperatures, similar to those of modern coastal southern British Columbia roughly through Oregon. The southern British Columbia region underwent significant uplift in the Early Eocene (Ewing 1980), reflected in the contrast between the Okanagan Highlands and coeval warmer, subtropical-character forests in nearby lowland coastal regions (Rouse et al. 1971). The Okanagan Highlands climate has been determined by taxon-independent leaf physiognomy data from variables such as the size and shape of dicot leaves and by taxon-dependent analyses of the nearest-living-relative climatic associations of plant micro- and megafossils and insects (Figs. 2, 3; e.g. Rouse et al. 1971; Wolfe and Wehr 1987; Wolfe 1994; Wolfe et al. 1998; Archibald and Mathewes 2000; Greenwood et al. 2005; Dillhoff et al. 2005; Moss et al. 2005; Archibald 2007; Smith et al. 2009; Smith 2011). Paleobotanical indicators show mesic levels of precipitation.
10 Fine preservation of leaf compression fossils to the cellular level allows evaluation of other aspects of the paleoenvironment, including levels of atmospheric carbon dioxide (pCO2). Estimates of Early Eocene pCO2 vary widely, from near modern-day levels to an order of magnitude greater (Royer 2006; Smith et al. 2010). Resolving these is a key task in determining whether climate and pCO2 were then coupled. The stomatal frequency of fossil plants is one proxy measure that can be used to estimate paleo-pCO2, as the frequency of stomata on the leaf surface is regulated, at least in part, by levels of CO2 (Woodward 1987; Royer 2001). The ‘living fossil’ Ginkgo biloba is often used in the application of this proxy measure, as the fossil Ginkgo adiantoides is indistinguishable from the living species, and indicates that this lineage has persisted with little change in gross morphology since the Cretaceous (Royer et al. 2003). The exceptional preservation of Ginkgo with intact cuticle at the Okanagan Highlands Falkland locality (Fig. 5) has provided the opportunity to estimate pCO2 during the critical window of the Early Eocene. Stomatal counts on fossil Ginkgo leaves at Falkland indicate that pCO2 was >2x modern-day levels during the Early Eocene (Smith et al. 2010). Furthermore, evidence of cooling temperatures over time at Falkland is correlated with a trend toward decreasing pCO2, suggesting that they were coupled (Smith et al. 2010).
11 The Eocene was also a time of seasonally equable temperatures, with mild winters extending into high latitudes, unlike today when such low seasonality is most associated with low latitudes (Greenwood and Wing 1995; Zachos et al. 2008; Shellito et al. 2009).
COMMUNITIES: FAMILIAR AND STRANGE
12 The particular Okanagan Highlands mixture of temperate upland mean annual temperatures, and winters with few if any frost days, may have been of key importance in determining the distinctive character of its forests (Archibald and Farrell 2003; Greenwood et al. 2005). The paleoclimate appears to have allowed plant and insect taxa that are today associated with sub-tropical to tropical temperatures to co-exist with more typically temperate elements. Here, forests contained palms (Uhlia) and cycads, at some localities mixed with taxa such as spruce (Erwin and Stockey 1991; Greenwood et al. 2005).
13 Not only were these communities notable for their tropical–temperate mixture of organisms, but they possessed differing patterns of species richness than we would expect in modern temperate, mid-latitude forests. Diversity of insects (McAbee) and plants (Falkland, McAbee, Republic) has been shown to be surprisingly high; it is comparable to that found in modern tropical forests (Archibald et al. 2010; Smith et al. 2011), and the Falklands flora matches the hyperdiverse floras of Eocene South America (Wilf et al. 2005; Smith et al. 2011). Perhaps oddly, this tropical level of diversity seen in insects and plants is not reflected in fish.
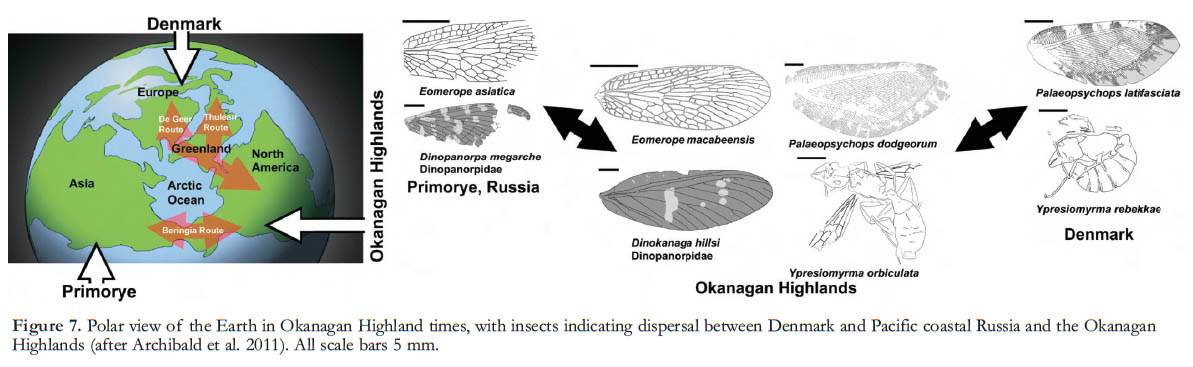
14 Some Okanagan Highlands plant and insect lineages are now native to distant regions such as Chile or Australia (e.g. Wing and Greenwood 1993; Archibald and Mathewes 2000; Archibald and Farrell 2003; Greenwood et al. 2005; Archibald et al. 2005b, 2006). In the Early Eocene, North America was connected to Europe across the Arctic via Greenland, and to East Asia via Beringia (Manchester 1999). Patterns of Okanagan Highlands insect and plant fossil occurrences have helped reveal the histories of particular lineages that have disjunct distributions in the modern world (e.g. DeVore et al. 2005; Archibald and Makarkin 2006; Archibald 2009; DeVore and Pigg 2007, 2010). The Okanagan Highlands forests show similarities to those of modern eastern USA, but also include many taxa that today occur in East Asia and the Pacific Northwest, alongside some that are now extinct, helping elucidate the origins and history of lineages characteristic today of microthermal broad-leaved deciduous and coniferous forests (Wolfe 1987; Manchester 1999). For example, taxa that are characteristic of the eastern hardwood forests of North America today and also represented in the Okanagan Highlands flora include elm (Ulmus, Fig. 3L–M), beech (Fagus), hazel (Corylus), hornbeam (Carpinus), birch (Betula), maple (Acer, Fig. 3R) and others. Relict taxa that are today restricted to East Asia, but were more widespread in the Early Eocene, include typical Okanagan Highlands taxa such as dawn redwood (Metasequoia, Fig. 3A), maidenhair tree (Ginkgo, Fig. 3B), and golden larch (Pseudolarix, Fig. 3F), as well as more rare occurrences of Dipteronia, Eucommia (Fig. 6C), Koelreuteria (Fig. 3Q), Tetracentron (Fig. 3E) and Photinia (Fig. 2P). Genera that are found today in the Pacific Northwest of North America, and that were also present during the Early Eocene, include cedar (Thuja), redwood (Sequoia), spruce (Picea), fir (Abies, Fig. 3D), pine (Pinus, Fig. 3C), alder (Alnus, Fig. 3J–K), cherry (Prunus, Fig. 3O) and others.
CONCLUSIONS
15 While Lagerstätten such as the Late Eocene Florissant and Middle Eocene Messel preserve an extraordinary range of life in fine detail (Meyer 2003; Schaal and Ziegler 1992), they reflect the environments around single lakes. The multiple Early Eocene deposits of the Okanagan Highlands, however, preserve a profusion of life surrounding a series of lakes spanning about 1000 kilometres north to south, providing an unprecedented opportunity to examine an extended transect of temperate uplands in a warm world. These fossils were deposited at a key moment in the emergence of modern terrestrial lineages and communities before the onset of our current icehouse world climate, an ancient world with a mixture of familiar and strange life forms and environments. The extraordinary Lagerstätten of the Okanagan Highlands offer an excep tional window into the formation of life in our modern world and constitute a unique comparative framework within which to examine the processes that govern its nature.
 Display large image of Figure 7
Display large image of Figure 7Funding for this research was provided by NSERC through Discovery Grants to DRG, JFB and RWM, a Canada Graduate Scholarship to RYS and a PostDoctoral Fellowship to SBA, and by facilities funded by a Canada Foundation for Innovation grant to DRG. We wish to acknowledge Richard Hebda of the Royal BC Museum for permission to collect at the BC sites, and John Howard and Scott McMillan of BC Parks for facilitating work at Driftwood Canyon Provincial Park (under BC Parks permit). We also acknowledge the generous assistance of Catherine Brown and Karl Volkman of the Stonerose Interpretive Center in Republic, WA. We thank Marlow Pellatt (Parts Canada) for access to microphotography equipment. We thank our colleagues for their advice and input. We thank David Rudkin and Graham Young for their efforts in organizing both the Lagerstätten symposium at GeoCanada 2010 in Calgary, Alberta, and this series, and further thank them, Mark Wilson, and Mike Wilson for their helpful comments on this manuscript.