Series
Geoscience of Climate and Energy 4.
Rapid Carbon Injection and Transient Global Warming during the Paleocene– Eocene Thermal Maximum
Appy SluijsInstitute of Environmental Biology, Laboratory of Palaeobotany and Palynology, Utrecht University, Utrecht, The Netherlands
a.sluijs@uu.nl
INTRODUCTION
1 The Paleocene–Eocene Thermal Maximum (PETM), ~55.5 My ago, was a geologically brief (~170 ky) episode of globally elevated temperature that was superimposed on the long-term Late Paleocene and Early Eocene warming trend (Fig. 1). It was marked by a 5-8°C warming in both low and high-latitude regions, a perturbation of the hydrological cycle and major biotic response on land and in the oceans, including radiations, extinctions and migrations (see overviews in Bowen et al. 2006 and Sluijs et al. 2007a). In addition, the PETM is associated with a pronounced negative carbon-isotope excursion (CIE), recorded as a >2.5‰ decrease in the stable carbon isotope composition (δ13C) of sedimentary components (e.g. Kennett and Stott 1991; Koch et al. 1992; Fig. 1). The CIE can only be explained by a carbon ‘burp’ – a massive (at least 1.5×1018 g or 1500 Gt) injection of 13C-depleted carbon into the ocean-atmosphere system (Dickens et al. 1995).
2 Recent work has focused on elucidating the injection mechanism(s) and the quantity of carbon that caused the CIE, but has also addressed the question of whether the 13C-depleted carbon caused the warming or acted as a positive feedback in an already warming world. Was the PETM a unique event in the early Paleogene greenhouse world, and what is the relevance of the PETM for current carbon injection from fossil fuel burning?
OCEAN ACIDIFICATION
3 Analogous to the modern situation, the injection of a large mass of CO2 or CH4 (which would have been oxidized to CO2 within a century at most; Schmidt and Shindell 2003) should have increased the acidity of the ocean. As a result, a shallowing of the calcite compensation depth (CCD) and dissolution of deep-sea carbonates should have occurred, thereby buffering the seawater pH change (Dickens et al. 1997). Indeed, the dissolution of deep-sea carbonates during the PETM has been documented in various deep-ocean basins, based on the occurrence of clay layers (the residual sedimentary component in the deep sea after calcite dissolution) as well as biogenic calcite fragmentation (e.g. Zachos et al. 2005; Fig. 1). The severity of dissolution, however, appears to have been highly variable between different basins, perhaps caused by spatial variability in bioturbation (Zeebe and Zachos 2007; Panchuk et al. 2008). Moreover, the magnitude of dissolution should have been equivalent to the amount of injected carbon and has, as such, been used in modelling experiments to elucidate the source and volume of injected carbon (Panchuk et al. 2008). According to that study, nearly 7000 Gt of carbon, derived from multiple sources, was injected during the PETM.
CARBON SEQUESTRATION
4 The distribution of deep-sea carbonate points to one mechanism of carbon sequestration during recovery of the PETM. Carbonate accumulation rates at many sites appear to have been very high in the latter stages of the PETM: the lysocline (the depth in the ocean below which the rate of dissolution of calcite increases dramatically) was located even deeper than before the PETM, resulting in the sequestration of large amounts of carbon (e.g. Kelly et al. 2005; Zachos et al. 2005). This phenomenon is model-predicted (Dickens et al. 1997) and was probably driven by silicate weathering, which slowly recharged the ocean with carbonate ions and eventually led to carbonate ion over-saturation and extremely good preservation of calcite on the sea floor. Another mechanism of carbon sequestration leading to the termination of the PETM comes from recent work that revealed a major increase in carbon burial rates on continental shelves during the event (Sluijs et al. 2008; John et al. 2008). At several sites, calcite burial rates increased (John et al. 2008), but increased organic carbon burial may have been significant as well. Particularly in the Arctic but also in the Tethys, organic carbon burial increased during the PETM due to increased river runoff, which caused increased organic production in the oceans, as well as stratification of the water column and bottom water anoxia (Sluijs et al. 2008). Excess burial perhaps amounted to roughly 800 Gt of carbon during the PETM in the Arctic Ocean alone (Sluijs et al. 2008).
Figure 1. A) Benthic foraminifer δ18O record from 62-48 My, adapted from Zachos et al. (2001); data for ETM2 represent the Cibicidoides record from Lourens et al. (2005). The δ18O values (green diamonds) represent corrected values (cf. Zachos et al. 2001). The red line is a 5 point moving average. An additional correction of -0.25 ‰ was applied to the ETM2 Cibicidoides values to synchronize baseline levels between the records. ETM2: Eocene Thermal Maximum 2 (also referred to as H1 or Elmo); PETM: Paleocene–Eocene Thermal Maximum; VPDB: Vienna Pee Dee Belemnite δ18O and δ13C standard. B) Compilation of δ13C and δ18O values across the PETM of planktonic foraminifera (surface dweller Acarinina and thermocline dweller Subbotina spp.; mostly single specimen), benthic foraminifera (Nuttallides truempyi) and bulk carbonate from ODP Site 690 at Maud Rise in the Weddell Sea (data compilation and figure modified from Sluijs et al. 2007a). BFE refers to the main phase of benthic foraminifer extinction. CIE: carbon-isotope excursion. C) Carbonate weight % record and core photo of ODP Site 1263 at Walvis Ridge, southeastern Atlantic (adapted from Zachos et al. 2005), showing dissolution and resulting clay layer. mbsf: metres below seafloor; mcd: metres composite depth.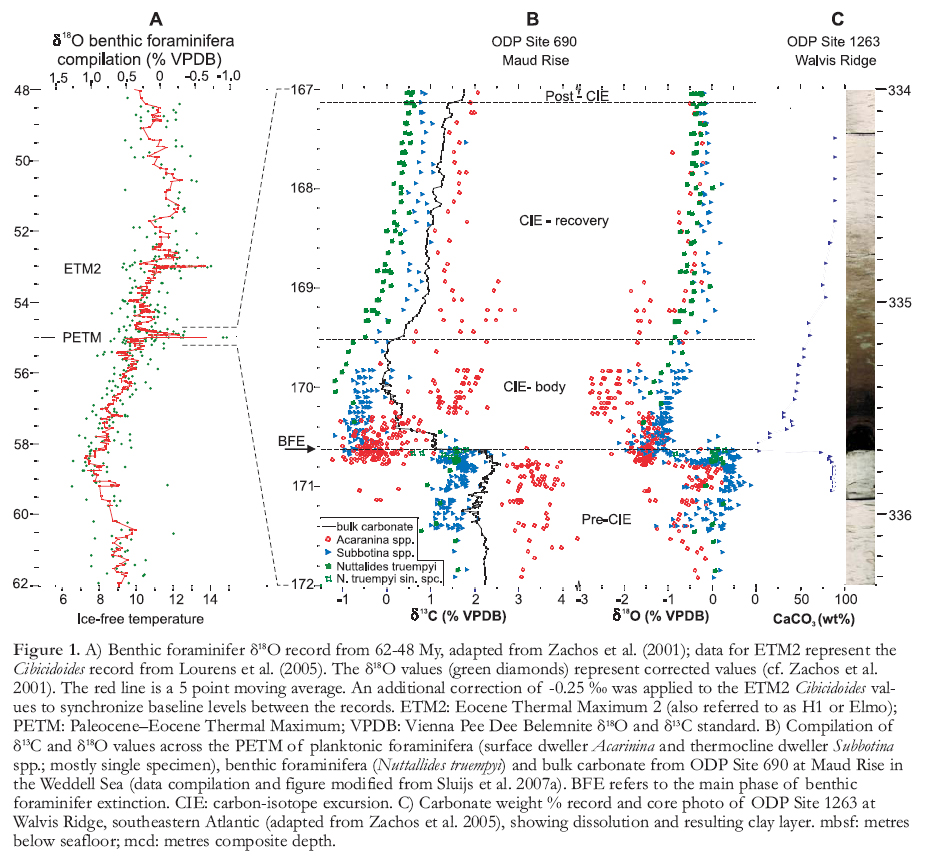
MERIDIONAL TEMPERATURE GRADIENTS
5 The application of the organic pale-othermometer TEX86 (based on the distribution of species of membrane lipids of the marine picoplankton Crenarchaeota; Schouten et al. 2002), as well as evidence from oxygen isotopes (Fig. 1) on well-preserved foraminifera, has recently led to a much better quantification of sea surface temperatures (SSTs) across the PETM. Prior to the PETM excursion, ‘background’ Late Paleocene–Early Eocene was already warm, with SSTs of ~32 °C in the tropics (Pearson et al. 2007), although the recently revised calibration of the TEX86 proxy (Kim et al. 2008) suggests that tropical temperatures were several degrees higher. Mid- and high-latitude surface ocean temperatures were approximately 25 °C and 17 °C, respectively (e.g. Sluijs et al. 2006, 2007b; Zachos et al. 2006). During the PETM, tropical as well as mid-latitude and Arctic SSTs rose by 5-8°C (Zachos et al. 2003, 2006; Sluijs et al. 2006; Fig. 2). Such temperatures in the high Arctic are supported by biogeographical data, such as the abundant occurrence of subtropical dinoflagellates (Sluijs et al. 2006), and other biogeochemical information (e.g. Weijers et al. 2007). Hence, meridional temperature gradients were significantly smaller than at present during both background and PETM conditions, although it remains unclear if the Arctic data represent mean annual or summer temperatures (Sluijs et al. 2006). Yet even if they represent summer temperatures and tropical temperatures higher than previously estimated (Pearson et al. 2007; Huber 2008), current-generation fully-coupled climate models overestimate Paleocene–Eocene meridional gradients, despite the model being fed with Eocene geography and high CO2 concentrations (Huber and Nof 2006; Fig. 2). This suggests that higher-than-modern greenhouse gas concentrations must have operated in conjunction with feedback mechanisms that either amplified polar temperatures or cooled the tropics, but that have not been incorporated in the models (Sluijs et al. 2006). Potential feedback mechanisms include polar stratospheric clouds (Sloan and Pollard 1998) and hurricane-induced ocean mixing (Emanuel et al. 2004; Sriver and Huber 2007), to account for polar warming and tropical cooling, respectively.
6 Interestingly, it appears that the meridional temperature gradient did not further decrease during the PETM. This can be partly explained by the absence of ice-albedo feedbacks, since the Arctic was already ice-free prior to the PETM. Additionally, it implies that the mechanism that caused the reduced meridional temperature gradient did not become amplified during the PETM (Sluijs et al. 2006).
ADDITIONAL EARLY EOCENE HYPERTHERMALS
7 Recent work shows that a similar warming phase occurred at ~53.5 My (referred to in the literature as H-1, Elmo or Eocene Thermal Maximum 2 (ETM2); Lourens et al. 2005), and possible additional phases at ~53.1 My (I-1) and ~52.3 My (K or X) (Cramer et al. 2003; Röhl et al. 2005; Nicolo et al. 2007). Although documentation of these phases is, as yet, relatively incomplete, the available information indicates that these hyperthermals are also associated with massive injection of 13C-depleted carbon, ocean acidification and perturbations of the hydrological cycle, although less pronounced than during the PETM. Orbital tuning of the complete Late Paleocene and Early Eocene record at Walvis Ridge (South Atlantic) has indicated a link between the timing of the hyperthermals and eccentricity maxima (Lourens et al. 2005; Westerhold et al. 2007), which would have implications for the mechanisms that caused global change during the hyperthermals.
LEADS AND LAGS AND MECHANISMS OF CARBON INPUT
8 One prominent example of biotic change associated with the PETM is recorded along continental margins, where sedimentary sequences across the event from all latitudes contain high abundances of dinoflagellate cysts belonging to the subtropical genus Apectodinium (Crouch et al. 2001; Sluijs et al. 2007a). In part, this must be associated with the PETM warming. However, in stratigraphically expanded marginal marine sections from the New Jersey Shelf and the North Sea, as well as a section in New Zealand, the onset of the Apectodinium acme started some 5 ky prior to the CIE (Sluijs et al. 2007b; Fig. 2). Additionally, the onset of the PETM SST warming at New Jersey appears to have led the CIE by several thousands of years (but lagged the onset of the Apectodinium acme; Sluijs et al. 2007b). This indicates that warm SST was not the only environmental control on Apectodinium abundances. Moreover, it suggests that the carbon ‘burp’ that caused the CIE was a result of initial climate change and acted as a positive feedback. This scenario fits a model that relates the CIE to CH4 release from submarine hydrates (Dickens et al. 1995). If this pre-CIE warming was global, it was likely induced by greenhouse forcing, suggesting that the PETM warming and ocean acidification were caused by at least two sources of carbon (Sluijs et al. 2007b).
Figure 2. High-resolution records across the onset of the PETM at Bass River, New Jersey Shelf Sites; redrawn from Sluijs et al. (2007b). Solid horizontal line at ~357.3 mbs represents the onset of the carbon-isotope excursion (CIE); dashed lines represent the onsets of the Apectodinium acme and surface warming. BC: bulk carbonate; DINO: dinocysts; VPDB: Vienna Pee Dee Belemnite δ13C standard; SST: sea-surface temperature; TEX86: Crenarcheota (marine picoplankton) paleothermometer; mbs: metres below surface.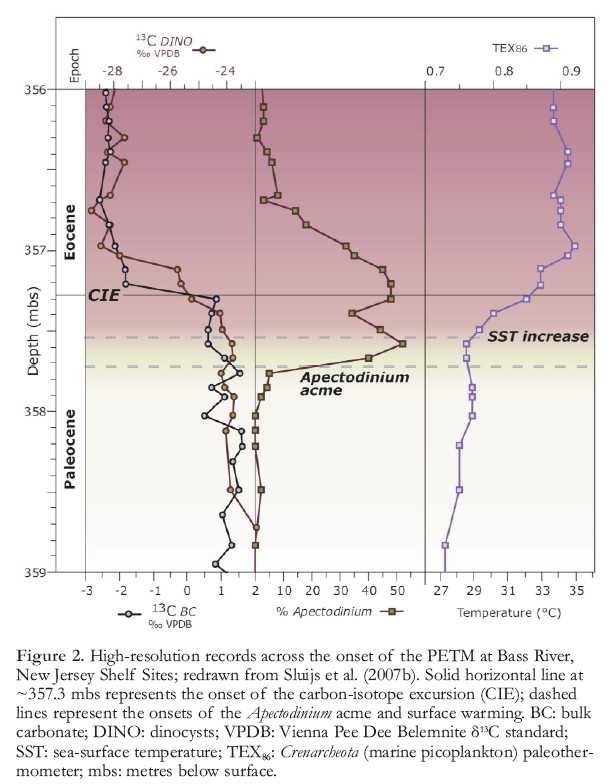
CONCLUDING REMARKS
9 The past years of research on the PETM and the newly discovered additional hyperthermals have resulted in a clearer picture of these critical phases in Earth’s history. Improved drilling techniques (Integrated Ocean Drilling Program) have resulted in the recovery of complete sections, and new analytical techniques have contributed to much better quantitative estimates of surface temperatures. Moreover, high-resolution studies on expanded marginal marine sequences have identified leads and lags in the interaction between the climate system and the carbon cycle during the onset of the PETM. The new data provide fundamental constraints for modelling global climate and carbon cycling and are increasingly leading to a much better description and understanding of the state and dynamics of a (in this case, early Paleogene) greenhouse world.
ACKNOWLEDGEMENTS
I thank the Netherlands Organisation for Scientific Research (NWO) for funding (VENI grant 863.07.001), Reg Wilson for editorial handling and Andrew Miall for inviting me to speak at the Gussow–Nuna Geoscience of Climate Change meeting, October 2008, Banff, Canada. I thank all ‘hyper-thermal’ colleagues, including Stefan Schouten, Jim Zachos, Jerry Dickens, Ursula Röhl, Lucas Lourens, Jaap Sinninghe Damsté, Gert-Jan Reichart, Isabella Raffi, Matthew Huber, Gabe Bowen, Mark Pagani, Ellen Thomas, Erica Crouch, Cedric John, Steven Bohaty, Hans Brumsack, Simone Gale-otti, Phil Gingerich, Chris Hollis, Clay Kelly, Dick Kroon, Lee Kump, Micah Nicolo, Rich Pancost, Paul Pearson, Domenico Rio, Stephen Schellenberg, Lisa Sloan, Francesca Smith, Lucy Stap, Heather Stoll, Debbie Thomas, Scott Wing, Richard Zeebe and the ones we forgot, for discussions over the past years. I also thank NWO for their continued support of the Integrated Ocean Drilling Program (IODP).REFERENCES
Bowen, G.J., Bralower, T.J., Delaney, M.L., Dickens, G.R., Kelly, D.C., Koch, P.L., Kump, L.R., Meng, J., Sloan, L.C., Thomas, E., Wing, S.L., and Zachos, J.C., 2006, Eocene hyperthermal event offers insight into greenhouse warming: EOS, Transactions of the American Geophysical Union, v. 87, p. 165-169.
Cramer, B.S., Wright, J.D., Kent, D.V., and Aubry, M.-P., 2003, Orbital climate forcing of δ13C excursions in the late Paleocene — early Eocene (chrons C24n–C25n): Paleoceanography, v. 18, doi: 10.1029/2003PA000909.
Crouch, E.M., Heilmann-Clausen, C., Brinkhuis, H., Morgans, H.E.G., Rogers, K.M., Egger, H., and Schmitz, B., 2001, Global dinoflagellate event associated with the late Paleocene thermal maximum: Geology, v. 29, p. 315-318.
Dickens, G.R., O’Neil, J.R., Rea, D.K., and Owen, R.M., 1995, Dissociation of oceanic methane hydrate as a cause of the carbon isotope excursion at the end of the Paleocene: Paleoceanography, v. 10, p. 965-971.
Dickens, G.R., Castillo, M.M., and Walker, J.C.G., 1997, A blast of gas in the latest Paleocene: Simulating first-order effects of massive dissociation of oceanic methane hydrate: Geology, v. 25, p. 259-262.
Emanuel, K., DesAutels, C., Holloway, C., and Korty, R., 2004, Environmental control of tropical cyclone intensity: Journal of the Atmospheric Sciences, v. 61, p. 843-858.
Huber, M., 2008. A Hotter Greenhouse? Science, v. 321, p. 353-354, doi: 10.1126/science.1161170
Huber, M., and Nof, D., 2006, The ocean circulation in the southern hemisphere and its climatic impacts in the Eocene: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 231, p. 9-28.
John, C.M., Bohaty, S.M., Zachos, J.C., Sluijs, A., Gibbs, S.J., Brinkhuis, H., and Bralower, T.J., 2008, North American continental margin records of the Paleocene-Eocene thermal maximum: Implications for global carbon and hydrological cycling: Paleoceanography, v. 23, PA2217, doi:2210.1029/2007PA001465.
Kelly, D.C., Zachos, J.C., Bralower, T.J., and Schellenberg, S.A., 2005, Enhanced terrestrial weathering/runoff and surface ocean carbonate production during the recovery stages of the Pale-ocene-Eocene thermal maximum: Paleoceanography, v. 20, PA4023, doi:10.1029/2005PA001163.
Kennett, J.P., and Stott, L.D., 1991, Abrupt deep-sea warming, palaeoceanographic changes and benthic extinctions at the end of the Palaeocene: Nature, v. 353, p. 225-229.
Kim J.-H., Schouten S., Hopmans E. C., Donner B., and Sinninghe Damsté J. S. (2008) Global sediment core-top calibration of the TEX86 paleothermometer in the ocean. Geochim. Cosmochim. Acta, v. 72, p. 1154-1173.
Koch, P.L., Zachos, J.C., and Gingerich, P.D., 1992, Correlation between isotope records in marine and continental carbon reservoirs near the Palaeocene/Eocene boundary: Nature, v. 358, p. 319-322.
Lourens, L.J., Sluijs, A., Kroon, D., Zachos, J.C., Thomas, E., Röhl, U., Bowles, J., and Raffi, I., 2005, Astronomical pacing of late Palaeocene to early Eocene global warming events: Nature, v. 435, p. 1083-1087.
Nicolo, M.J., Dickenc, G.R., Hollis, C.J., and Zachos, J.C., 2007, Multiple early Eocene hyperthermals: Their sedimentary expression on the New Zealand continental margin and in the deep sea: Geology, v. 35, p. 699-702.
Pearson, P.N., van Dongen, B.E., Nicholas, C.J., Pancost, R.D., Schouten, S., Singano, J.M., and Wade, B.S., 2007, Stable warm tropical climate through the Eocene Epoch: Geology, v. 35, p. 211-214.
Panchuk, K., Ridgwell, A., and Kump, L.R., 2008, Sedimentary response to Pale-ocene-Eocene Thermal Maximum carbon release: A model-data comparison: Geology, v. 36, p. 315-318.
Röhl, U., Westerhold, T., Monechi, S., Thomas, E., Zachos, J.C., and Donner, B., 2005, The third and final early Eocene thermal maximum: characteristics, timing, and mechanisms of the “X” event: Geological Society of America Annual Meeting – Abstracts, v. 37, p. 264.
Schmidt, G.A., and Schindell, D.T., 2003, Atmospheric composition, radiative forcing, and climate change as a consequence of a massive methane release from gas hydrates: Paleoceanography, v. 18, doi:10.1029/2002PA000757.
Schouten, S., Hopmans, E.C., Schefuß, E., and Sinninghe Damsté, J.S., 2002, Distributional variations in marine crenarchaeotal membrane lipids: A new tool for reconstructing ancient sea water temperatures?: Earth and Planetary Science Letters, v. 204, p. 265-274.
Sloan, L.C., and Pollard, D., 1998, Polar stratospheric clouds: A high latitude warming mechanism in an ancient greenhouse world: Geophysical Research Letters, v. 25, p. 3517-3520.
Sluijs, A., Schouten, S., Pagani, M., Woltering, M., Brinkjuis, H., Sinninghe Damsté, J.S., Dickens, G.R., Huber, M., Reichart, G.-J., Stein, R., Matthiessen, J., Lourens, L.J., Pedentchouk, N., Backman, J., Moran, K., and the Expedition 302 Scientists, 2006, Subtropical Arctic Ocean temperatures during the Palaeocene/Eocene thermal maximum: Nature, v. 441, p. 610-613.
Sluijs, A., Bowen, G.J., Brinkhuis, H., Lourens, L.J., and Thomas, E., 2007a, The Palaeocene-Eocene thermal maximum super greenhouse: Biotic and geochemical signatures, age models and mechanisms of global change, in: Williams, M., Hayward, A., Gregory, J., and Schmidt, D. (eds), Deep Time Perspectives on Climate Change: Marrying the Signal from Computer Models and Biological Proxies: The Micropalaeontological Society, Special Publications, The Geological Society, London, p 323-349.
Sluijs, A., Brinkhuis, H., Schouten, S., Bohaty, S.M., John, C.M., Zachos, J.C., Sinninghe Damsté, J.S., Crouch, E.M., and Dickens, G.R., 2007b, Environmental precursors to rapid light carbon injection at the Paleocene/Eocene boundary: Nature, v. 450, p. 1218-1221.
Sluijs, A., Röhl, U., Schouten, S., Brumsack, H.-J., Sangiorgi, F., Sinninghe Damsté, J.S., and Brinkhuis, H., 2008, Arctic late Paleocene – Early Eocene pale-oenvironments with special emphasis on the Paleocene – Eocene thermal maximum (Lomonosov Ridge, IODP Expedition 302): Paleoceanography, v. 23, PA1S11, doi:10.1029/2007PA001495.
Sriver, R.L., and Huber, M., 2007, Observational evidence for an ocean heat pump induced by tropical cyclones: Nature, v. 447, p. 577-580.
Weijers, J.W.H., Schouten, S., Sluijs, A., Brinkhuis, H., and Sinninghe Damste, J.S., 2007, Warm arctic continents during the Palaeocene-Eocene thermal maximum: Earth and Planetary Science Letters, v. 261, p. 230-238.
Westerhold, T., Röhl, U., Laskar, J., Raffi, I., Bowles, J., Lourens, L.J., and Zachos, J.C., 2007, On the duration of Magnetochrons C24r and C25n, and the timing of early Eocene global warming events: Implications from the ODP Leg 208 Walvis Ridge depth transect: Paleoceanography, v. 22, PA2201, doi:10.1029/2006PA001322.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K., 2001, Trends, rhythms, and aberrations in global climate 65 Ma to present: Science, v. 292, p. 686-693.
Zachos, J.C., Wara, M.W., Bohaty, S., Delaney, M.L., Petrizzo, M.R., Brill, A., Bralower, T.J., and Premoli Silva, I., 2003, A transient rise in tropical sea surface temperature during the Pale-ocene-Eocene thermal maximum, Science, v. 302, p. 1551-1554.
Zachos, J.C. Röhl, U., Schellenberg, S.A., Sluijs, A., Hodell, D.A., Kelly, D.C., Thomas, E., Nicolo, M., Raffi, I., Lourens, L.J., McCarren, H., and Kroon, D., 2005, Rapid acidification of the ocean during the Paleocene-Eocene Thermal Maximum: Science, v. 308, p. 1611-1615.
Zachos, J.C., Schouten, S., Bohaty, S., Quattlebaum, T., Sluijs, A., Brinkhuis, H., Gibbs, S., and Bralower, T.J., 2006, Extreme warming of mid-latitude coastal ocean during the Paleocene-Eocene Thermal Maximum: Inferences from TEX86 and Isotope Data: Geology, v. 34, p. 737-740.
Zeebe, R.E., and Zachos, J.C., 2007, Reversed deep-sea carbonate ion basin gradient during the Paleocene-Eocene thermal maximum: Paleoceanography, v. 22, PA3301, doi:10.1029/2006PA001395.